 Mixing
Mixing
獸腳亞目
| 兽脚亚目 化石时期:三疊紀晚期至今 231.4–0 Ma PreЄ Є O S D C P T J K Pg N | |
|---|---|
 | |
| 最先被分类为兽脚亚目的异特龙 | |
科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 爬行纲 Reptilia |
| 演化支: | 恐龙总目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 演化支: | 真蜥臀类 Eusaurischia |
| 亚目: | 兽脚亚目 Theropoda Marsh, 1881 |
| 演化支 | |
| |
獸腳亞目(学名:Theropoda)屬於蜥臀目,是群雙足恐龍。雖然牠們主要是肉食性動物,但是在白堊紀時期,一部份的獸腳類恐龍演變成為草食性、雜食性、甚至食蟲動物。獸腳類最早出現在三疊紀晚期的卡尼階,大約是2億3000萬年前。牠們從侏儸紀早期開始成為獨占廣大陸地的肉食動物,直到6500萬年前的白堊紀結束為止。獸腳亞目也是恐龍總目中唯一的肉食族群,其餘的鳥臀目所有恐龍、以及和獸腳亞目同為蜥臀目的蜥腳亞目都已經改為素食,但他們的祖先卻肉食素食都有。
獸腳亞目後來演變出一類特殊的小型恐龍,稱為虛骨龍類,虛骨龍經過幾千萬年的演化可能演變成了9,300個物種的鳥類的共同祖先。而目前已知最早的鳥類在侏儸紀晚期已經出現,例如始祖鳥,並且在恐龍還存活的白堊紀就已經有例如黃昏鳥和孔子鳥之類的現代鳥類生存了。獸腳亞目與鳥類之間的共同點包括:腳掌有三趾(牠們的第一趾其實萎縮在腳部後方)、胸部上的「叉骨」、與充滿空氣的骨頭。此外,某些獸腳類恐龍的手臂擁有羽毛(但不應具備飛行能力,例如恐爪龍和盜伶龍)和孵卵行為。
目录
1 演化歷程
2 古生物學
2.1 前肢功能
2.2 鱗片、羽毛
2.3 食性
3 分類學
3.1 分類歷史
3.2 種系發生學
4 體型
5 參考資料
演化歷程
在三疊紀晚期,一些原始獸腳類與獸腳類恐龍便已經出現。最早且最為原始的肉食性恐龍是阿根廷的始盜龍與艾雷拉龍下目。艾雷拉龍下目最早出現於三疊紀的卡尼階後期到諾利階(Norian)早期。在北美洲與南美洲可以發現這類恐龍化石,此外牠們可能也出現在印度與非洲南部。艾雷拉龍下目同時擁有一些較為原始以及較為晚近的恐龍特徵,是一種鑲嵌演化(Mosaic evolution;不同特徵擁有不同演化速率)的現象。古生物學家們本來認為艾雷拉龍類是獸腳類的物種之一,不過後來發現牠們可能是原始的蜥臀目恐龍,甚至可能早於蜥臀目與鳥臀目的分化。根據2009年新發現的太陽神龍,顯示艾雷拉龍類可能是群早期獸腳類恐龍[1]。

獸腳亞目的足跡
最早且最原始的獸腳亞目恐龍(或者說是真獸腳類)為腔骨龍超科。腔骨龍超科是群輕型、分布廣泛的群居恐龍。腔骨龍科包含小型掠食動物(例如腔骨龍、合踝龍),以及大型掠食動物(例如身長6公尺的雙脊龍)。這些成功的動物生存於晚三疊紀的晚卡尼階到早侏儸紀的托阿爾階。但在早期的親緣分支分類法研究裡,腔骨龍超科被分類於角鼻龍下目,並且被認為是更先進獸腳類的一個旁支[2];目前腔骨龍超科被認為可能是其他獸腳類的祖先,如此一來,牠們將成為並系群集合[3][4]。
較為先進的角鼻龍下目(包括角鼻龍與食肉牛龍)生存於勞亞大陸,牠們出現於早侏儸紀,並存活到晚侏儸紀。角鼻龍下目與更先進的堅尾龍類近親同時競爭,其中的阿貝力龍超科支系,在岡瓦納大陸存活到白堊紀末期。
堅尾龍類比角鼻龍下目更為特化。牠們可分為兩個演化支:較原始的斑龍超科(或稱棘龍超科、蠻龍超科)、以及較先進的鳥獸腳類。斑龍超科在中侏儸紀到早白堊紀最為普遍,其中的棘龍科存活到中白堊紀。鳥獸腳類更為接近鳥類,並且還可分為兩個演化支:肉食龍下目(例如異特龍)、虛骨龍類。虛骨龍類是一個非常大群且多樣化的演化支,在白堊紀時期非常普遍。
因此,在侏儸紀晚期,有四個獸腳亞目支系存活者,包括:角鼻龍下目、斑龍超科、肉食龍下目、以及虛骨龍類,牠們以豐富的小型與大型草食性恐龍為食。這四個演化支都存活到白堊紀,其中只有阿貝力龍超科、異特龍超科與虛骨龍類存活到白堊紀末;而阿貝力龍超科、異特龍超科分布於岡瓦納大陸,虛骨龍類主要分布於勞亞大陸。
在所有獸腳亞目演化支中,虛骨龍類是最多樣化的一個演化支。有些虛骨龍類演化支繁盛於白堊紀,例如:暴龍超科(包含暴龍)、馳龍科(包含迅猛龍與恐爪龍,牠們類似已知最古老的鳥類:始祖鳥)[5][6]、傷齒龍科、雜食性的偷蛋龍下目、草食性的似鳥龍科與鐮刀龍超科、以及鳥翼類[7]。這些不同的演化支起源於侏儸紀晚期,甚至是侏儸紀中期,但在白堊紀早期才開始繁盛起來。少數古生物學家,例如葛瑞格利·保羅(Gregory S. Paul),提出在這些先進獸腳亞目中,某些或者全部其實是從會飛行的恐龍,或者是類似始祖鳥的原始鳥類演化而來,並失去了牠們的飛行能力,重新成為陸地動物[8]。需要更多的化石證據才能了解這些先進獸腳亞目恐龍之間的正確關係。
古生物學
前肢功能
與人類、其他靈長目相比,獸腳類恐龍的前肢活動範圍不大,與大眾媒體與讀物中的形象相反[9]。獸腳類與原蜥腳類恐龍無法前掌前後轉動牠們的手掌,手掌只能朝下、或朝後。人類的橈骨與尺骨關節,允許他們隨意轉動手掌。在蜥臀目恐龍的肘部關節中,橈骨末端接至尺骨的溝,妨礙大範圍的動作。同樣地,其腕部關節的活動方向、活動範圍也有限[10]。
某些物種的手掌與手指可以做出較靈活的動作,例如肉食龍下目的高棘龍、以及原始的艾雷拉龍下目、雙脊龍科。虛骨龍類的前肢開始出現功能的轉變,肩膀關節節相當靈活,可將前肢平舉;可飛行鳥類的前肢運動達到極限。但是,似鳥龍下目與馳龍科的手掌、手指較不靈活。手盜龍類(包含馳龍類)的手腕有半月形骨頭,使腕部關節相當靈活,可以做出類似鳥類的手掌收疊於前肢側的動作[10]。
鱗片、羽毛
根據已發現的表皮痕跡,獸腳類恐龍的皮膚、覆蓋物相當多樣化。原始獸腳類恐龍具有小型、瘤狀鱗片。某些物種更具有由皮內成骨構成的大型鱗片,例如食肉牛龍[11]。
雖然原始鳥臀目曾被發現有毛狀結構,例如天宇龍[12];獸腳亞目的有羽毛恐龍僅限於虛骨龍類。美頜龍科與原始暴龍超科具有原始羽毛,可能是構造簡易的中空細毛[13]。鐮刀龍類的北票龍被發現有兩種型態的羽毛,分別是具有大型羽幹的羽毛,另一種則是結構簡單的原始羽毛。
大部分的有羽毛獸腳類恐龍,包含現代鳥類在內,部分身體仍保有鱗片。某些物種則同時保有鱗片、羽毛。擅攀鳥龍的基部尾巴仍保有爬行動物的鱗片[14]。侏羅獵龍的尾巴基部與後肢,發現了鱗片痕跡,化石周圍則發現疑似原始羽毛的可疑痕跡[15]。某些物種則全身幾乎覆蓋者羽毛,利如傷齒龍科的近鳥龍,其腳踝、腳掌也具有羽毛[16]。
食性
長久以來,獸腳類恐龍被認為全是肉食性恐龍;實際上,獸腳類恐龍的食物來源相當多樣化[17]。在19世紀到20世紀前半期,古生物學家根據獸腳類的銳利、有鋸齒邊緣的牙齒,推測牠們善於撕裂肉體。某些物種甚至被發現有獵食的習性,舉例而言,一個美頜龍標本的胃部發現一個蜥蜴化石,而一個迅猛龍標本則被發現正與原角龍發生打鬥。
第一群被確定不是肉食性的獸腳類恐龍是鐮刀龍超科(原名慢龍下目)。鐮刀龍類最初被認為屬於原蜥腳下目,具有小型頭部、喙狀嘴、針狀牙齒,是群草食性動物,其大型腹部可以容納較長的消化器官,詳細地消化植物纖維。直到近年,鐮刀龍類被發現是群高度特化的手盜龍類。近年的種系發生學研究也助於釐清不同食性的手盜龍類的演化關係。原始手盜龍類是雜食性動物,後來演化出不同的食性,鐮刀龍類、原始偷蛋龍類、少數鳥翼類演化成草食性動物,馳龍科演化成肉食性動物,阿瓦拉慈龍科、多數鳥翼類則是食蟲動物,而傷齒龍科、衍化的偷蛋龍類則屬於雜食性動物。除此之外,似鳥龍下目也是雜食性或草食性恐龍。棘龍科與某些獸腳類恐龍(例如惡龍)更特化成以魚類為主食[18][19]。
分類學
分類歷史
獸腳亞目是由奧塞內爾·查利斯·馬什在1881年建立[20]。馬什當初建立獸腳亞目,只包含異特龍科,後來獸腳亞目被提升到獸腳目,並包含更多的肉食性恐龍的科,例如:斑龍科、美頜龍科、似鳥龍科、板龍科、近蜥龍科、以及Hallopodidae科(該科目前被認為是鱷魚的近親)。由於馬什將獸腳亞目提升到獸腳目,該分類單元取代了愛德華·德林克·科普在1866年先建立的Goniopoda(彎曲的足部);科普是馬什的競爭對手,而科普建立Goniopoda以包含肉食性的恐龍[21]。
在20世紀早期,部分古生物學家不再認為肉食性恐龍是個天然群體,例如休尼(Friedrich von Huene)。休尼不使用獸腳目,改使用哈利·絲萊(Harry Seeley)建立的蜥臀目,休尼將蜥臀目分為虛骨龍亞目與Pachypodosauria亞目。休尼將大部分小型獸腳類恐龍歸類於虛骨龍亞目,大型獸腳類恐龍與原蜥腳類則歸類於Pachypodosauria亞目;休尼認為Pachypodosauria亞目是蜥腳類恐龍的祖先(在當時,原蜥腳類恐龍被認為是肉食性,因為勞氏鱷目巨齒龍的頭骨與埃弗拉士龍的身體被拼湊在一起)[21]。在1922年,W.D. Matthew與巴納姆·布郎(Barnum Brown)敘述了第一種馳龍科(艾伯塔馳龍)[22],並首次將原蜥腳類排除於肉食性恐龍之外,並再度採用科普的Goniopoda,但當時不被普遍接受[21]。
在1956年以前,獸腳亞目曾長時間不被使用,直到阿爾弗雷德·羅默將蜥臀目重新分類為獸腳亞目與蜥腳亞目(Sauropoda),獸腳亞目可分為原蜥腳下目、虛骨龍下目、肉食龍下目;目前的分類法大致上遵循者羅默的版本。在1969年,恐爪龍與恐手龍的發現,衝擊了虛骨龍下目與肉食龍下目的二分法,因為牠們無法份類於任一個下目。在1970年代晚期,瑞欽·巴思缽(Rinchen Barsbold)將獸腳亞目改分類為:虛骨龍下目、恐爪龍下目、偷蛋龍下目、肉食龍下目、似鳥龍下目、恐手龍下目(Deinocheirosauria)[21]。
從1980年代開始,隨者親緣分支分類法與系統發生學的盛行,獸腳類恐龍的分類與演化關係逐漸明朗。在1986年,嘉克斯·高斯特(Jacques Gauthier)建立堅尾龍類,將獸腳亞目分為基礎型的角鼻龍下目與較衍化的堅尾龍類。同年,高斯特並建立了手盜龍類,以包含最類似鳥類的獸腳類恐龍。隨者近年許多關於獸腳類與鳥類關係的發現,大部分科學家認為鳥類是手盜龍類的直系後代,使得鳥類成為白堊紀-第三紀滅絕事件後,唯一倖存至今的恐龍[21]。
種系發生學
以下演化樹是從大衛·威顯穆沛(David Weishampel)等人2004年的《The Dinosauria》一書更改而來[23]。當中腔骨龍超科被歸類於角鼻龍下目,但近年的研究多將兩者列為分開的演化支。
| 獸腳亞目 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
體型
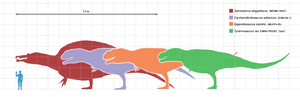
數種巨型獸腳類恐龍的體型相比。從左到右為:棘龍、鯊齒龍、南方巨獸龍、暴龍
過去數十年來,對一般大眾而言,暴龍是最大型的也是最著名的獸腳亞目恐龍。然而,自從暴龍被發現以後,已經有許多的其他大型獸腳亞目恐龍被敘述,包括:棘龍、鯊齒龍、南方巨獸龍、以及馬普龍[24]。在電影《侏儸紀公園III》中,棘龍被敘述成體型大於暴龍;而根據棘龍的原始標本與2006年的新發現化石,也顯示棘龍身長比暴龍長6公尺,而體重比暴龍重1公噸[25]。目前還沒有科學上的解釋,為何這些掠食動物成長的越來越大。
若不計算鳥類,已知最小型的獸腳類恐龍是耀龍,成年耀龍的身長約25公分,體重約164公克[26]。若計算現代鳥類,最小型的獸腳類恐龍是吸蜜蜂鳥,身長只有5.5公分,體重為1.9公克[27]。
參考資料
维基共享资源中相关的多媒体资源:獸腳亞目 |
维基物种中的分类信息:獸腳亞目 |
^ Nesbitt, S. J.; Smith, N. D.; Irmis, R. B.; Turner, A. H.; Downs, A. & Norell, M. A., A complete skeleton of a Late Triassic saurischian and the early evolution of dinosaurs, Science, 2009, 326 (5959): 1530–1533, doi:10.1126/science.1180350 .
^ Rowe, T., and Gauthier, J., (1990). "Ceratosauria." Pp. 151-168 in Weishampel, D. B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, University of California Press, Berkley, Los Angeles, Oxford.
^ Mortimer, M. (2001). "Rauhut's Thesis", Dinosaur Mailing List Archives, 4 Jul 2001.
^ Carrano, M. T., Sampson, S. D. and Forster, C. A., (2002). "The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar." Journal of Vertebrate Paleontology, 22(3): 510-534.
^ Ostrom, J.H. (1969). "Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana." Peabody Museum Natural History Bulletin, 30: 1-165.
^ Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster Co. (ISBN 0-671-61946-2)
^ Dingus, L. and Rowe, T. (1998). The Mistaken Extinction: Dinosaur Evolution and the Origin of Birds. Freeman.
^ Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. 472 pp. (ISBN 0-8018-6763-0)
^ Carpenter, K. (2002). "Forelimb biomechanics of nonavian theropod dinosaurs in predation." Senckenbergiana lethaea, 82(1): 59-76.
^ 10.010.1 Senter, P. and Robins, J.H. (2005). "Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour." Journal of Zoology, London, 266: 307–318.
^ Bonaparte, Novas, and Coria (1990). "Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia." Contributions in Science (Natural History Museum of Los Angeles County), 416: 41 pp.
^ Zheng, Xiao-Ting; You, Hai-Lu; Xu, Xing; Dong, Zhi-Ming. An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures. Nature. 19 March 2009, 458 (7236): 333–336. doi:10.1038/nature07856. 引文使用过时参数coauthors (帮助)
^ Göhlich, U.B., and Chiappe, L.M. (2006). "A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago." Nature, 440: 329-332.
^ Czerkas, S.A., and Yuan, C. (2002). "An arboreal maniraptoran from northeast China." Pp. 63-95 in Czerkas, S.J. (Ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1. The Dinosaur Museum, Blanding, U.S.A. PDF abridged version
^ Goehlich, U.B., Tischlinger, H., and Chiappe, L.M. (2006). "Juraventaor starki (Reptilia, Theropoda) ein nuer Raubdinosaurier aus dem Oberjura der Suedlichen Frankenalb (Sueddeutschland): Skelettanatomie und Wiechteilbefunde." Archaeopteryx, 24: 1-26.
^ Xu, X.; Zhao, Q.; Norell, M.; Sullivan, C.; Hone, D.; Erickson, G.; Wang, X.; Han, F. & Guo, Y. A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin. Chinese Science Bulletin. 2009, 54: 430–435. doi:10.1007/s11434-009-0009-6. Abstract
^ Zanno, L.E., Gillette, D.D., Albright, L.B., and Titus, A.L. (2009). "A new North American therizinosaurid and the role of herbivory in 'predatory' dinosaur evolution." Proceedings of the Royal Society B, Published online before print July 15, 2009, doi: 10.1098/rspb.2009.1029.
^ Longrich, N.R.; and Currie, P.J. Albertonykus borealis, a new alvarezsaur (Dinosauria: Theropoda) from the Early Maastrichtian of Alberta, Canada: Implications for the systematics and ecology of the Alvarezsauridae. Cretaceous Research. 2008,. online preprint: 239. doi:10.1016/j.cretres.2008.07.005. 引文使用过时参数coauthors (帮助)
^ Holtz, T.R., Jr., Brinkman, D.L., and Chandler, C.L. (1998). "Dental morphometrics and a possibly omnivorous feeding habit for the theropod dinosaur Troodon." GAIA, 15: 159-166.
^ Marsh, O.C. (1881). "Principal characters of American Jurassic dinosaurs. Part V." The American Journal of Science and Arts, series 3 21: 417-423.
^ 21.021.121.221.321.4 Rauhut, O.W. (2003). The Interrelationships and Evolution of Basal Theropod Dinosaurs. Blackwell Publishing, 213 pp. ISBN 090170279X
^ Matthew, W. D., and Brown, B. (1922) "The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta." Bulletin of the American Museum of Natural History, 46: 367-385.
^ Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.) (2004). The Dinosauria, Second Edition. University of California Press., 861 pp.
^ Therrien, F.; and Henderson, D.M. My theropod is bigger than yours...or not: estimating body size from skull length in theropods. Journal of Vertebrate Paleontology. 2007, 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2. 引文使用过时参数coauthors (帮助)
^ dal Sasso, C.; Maganuco, S.; Buffetaut, E.; and Mendez, M.A. New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities. Journal of Vertebrate Paleontology. 2005, 25 (4): 888–896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. 引文使用过时参数coauthors (帮助)
^ Zhang, Fucheng; Zhou, Zhonghe; Xu, Xing; Wang, Xiaolin and Sullivan, Corwin. A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers (455). Nature: 1105–1108. 23 October 2008. doi:10.1038/nature07447.
^ Conservation International (Content Partner); Mark McGinley (Topic Editor). 2008. "Biological diversity in the Caribbean Islands." In: Encyclopedia of Earth. Eds. Cutler J. Cleveland (Washington, D.C.: Environmental Information Coalition, National Council for Science and the Environment). First published in the Encyclopedia of Earth May 3, 2007; Last revised August 22, 2008; Retrieved November 9, 2009 [1].
| |||||||||||||||||||||||||||||||||||||||||||




Comments
Post a Comment