 Mixing
Mixing
二叠纪-三叠纪灭绝事件
二疊紀-三疊紀滅絕事件(Permian–Triassic extinction event,簡稱P-Tr)是一個大規模物种滅絕事件,發生於古生代二疊紀與中生代三疊紀之間,距今大約2億5140萬年[1][2]。若以消失的物種來計算,當時地球上70%的陸生脊椎動物,以及高達96%的海中生物消失[3];這次滅絕事件也造成昆蟲的唯一一次大量滅絕。計有57%的科與83%的屬消失[4][5]。在滅絕事件之後,陸地與海洋的生態圈花了數百萬年才完全恢復,比其他大型滅絕事件的恢復時間更長久[3]。此次滅絕事件是地質年代的五次大型滅絕事件中,規模最龐大的一次,因此又被正式稱為大死亡(The Great Dying)[6],或是大規模滅絕之母(Mother of all mass extinctions)[7]。
二疊紀-三疊紀滅絕事件的過程與成因仍在爭議中[8]。根據不同的研究,這次滅絕事件可分為一[1]到三[9]個階段。第一個小型高峰可能因為環境的逐漸改變,原因可能是海平面改變、海洋缺氧、盤古大陸形成引起的乾旱氣候;而後來的高峰則是迅速、劇烈的,原因可能是撞擊事件、超級火山爆發[10][11]、或是海平面驟變,引起甲烷水合物的大量釋放[12]。持续时间大约6万年[13]。
目录
1 年代測定
2 滅絕模式
2.1 海中生物
2.2 陸地無脊椎動物
2.3 陸地植物
2.3.1 植物生態系統
2.3.2 煤層缺口
2.4 陸地脊椎動物
2.5 滅絕模式的可能解釋
3 生態系統的復原
3.1 海洋生態系統的改變
3.2 陸地脊椎動物
4 可能的滅絕原因
4.1 陨石撞擊事件
4.2 火山爆發
4.3 甲烷水合物的氣化
4.4 海平面改變
4.5 海洋缺氧
4.6 硫化氫
4.7 盤古大陸的形成
4.8 多重原因
5 参考文献
6 延伸阅读
7 外部連結
年代測定
在西元二十世紀之前,二疊紀與三疊紀交界的地層很少被發現,因此科學家們很難準確地估算滅絕事件的年代與經歷時間,以及影響的地理範圍[14]。在1998年,科學家研究浙江省長興縣煤山附近的二疊紀/三疊紀岩層,他們採用鈾-鉛測年方法,研究鋯石中的鈾/鉛比例[2],估計二疊紀-三疊紀滅絕事件的發生年代為2億5140萬年前(誤差值為30萬年),並發現此後有隨着時間持續增高的滅絕比例[1]。在滅絕事件發生時,全球各地的碳13/碳12比例極速下降約9‰[15][16][17][18][19]。因為二疊紀/三疊紀的界線難以用放射性定年法測定,科學家們多用急遽下降的碳13/碳12比例,測定岩層中的二疊紀/三疊紀交界[20]。
在二疊紀與三疊紀交界時期,陸地與海生真菌曾有急遽的增加,原因應是當時有大量的植物與動物死亡,使得真菌得以迅速繁衍[21]。因為二疊紀/三疊紀的界線難以用放射性定年法測定,也缺乏參考用的標準化石,古動物學界一度使用真菌的繁衍高峰,測定岩層中的二疊紀/三疊紀交界。但是真菌定年法的最初提倡者,指出真菌的繁衍高峰,可能是三疊紀最早期生態系統的反覆發生現象[21]。真菌的繁衍高峰理論在最近幾年遭到許多質疑,例如:最常發現的真菌孢是Reduviasporonites,在近年被發現其實是藻類化石、[22][15]、真菌的繁衍高峰似乎不是全球性現象[23][24]、在許多地層中,真菌的繁衍高峰與二疊紀/三疊紀界線並不一致[25]。藻類被誤認為真菌胞,可能代表三疊紀時期全球各地湖泊增加,而非生物的大量死亡與腐爛[26]。在2009年的一個研究,則以生物化學方式分析Reduviasporonites的組成,例如:碳同位素、氮同位素、碳/氮比例,發現Reduviasporonites的組成接近真菌[27]。
關於這次滅絕的過程、經歷時間、以及不同生物群的滅絕模式,仍然相當模糊。一些證據顯示這次滅絕事件持續約數百萬年,而在二疊紀的最後100萬年,急遽地達到高峰[25][28]。在2000年,科學家研究浙江煤山的許多富含化石的地層,藉由統計分析指出,這次滅絕事件只有一個滅亡高峰期[1]。但是,之後近年的研究指出不同的生物群在不同的時間點滅亡;舉例而言,雖然年代難以確定,介形綱(Ostracoda)與腕足動物的大規模滅絕,先後相差了72萬年到122萬年之間[29]。在格陵蘭東部的一個化石保存良好的地層,動物群的衰退集中在6萬到1萬年之間,而植物群的衰退則持續約數十萬年之久,但也顯示滅絕事件造成全面性的影響[30]。一個較早期的研究,認為當時有兩個滅絕高峰期,相隔500萬年,這段期間的生物滅絕高於正常標準;而最後的滅絕高峰,造成當時約80%的海洋生物滅亡,其餘海生生物則多在第一次的滅絕高峰與滅絕間期消失;近年仍有研究引用這個假設[31]。若根據此一理論,第一次的滅絕高峰發生於二疊紀瓜達鹿白階末期[32]。
瓜達鹿白階末期滅絕事件對海生生物的衝擊,隨者地區與物種的不同,而有不同程度的影響。舉例而言,腕足動物與珊瑚有嚴重的大量滅亡[33][34]。所有的恐頭獸類滅亡,僅有一屬繼續存活[35];在有孔蟲門中,體型大型的紡錘蟲目(Fusulinida)也僅有費伯克蜓科(Verbeekinidae)繼續存活[36]。
滅絕模式
| 滅絕的海生動物 | 滅絕屬比例 | 備註 | |
|---|---|---|---|
海生無脊椎動物 | |||
有孔蟲門 | 97% | 紡錘蟲科滅絕,大部分在事件發生前滅絕 | |
放射蟲門(浮游生物) | 99%[37] | ||
珊瑚綱(海葵、珊瑚…等) | 96% | 四射珊瑚與床板珊瑚滅絕 | |
苔蘚動物門 | 79% | 小窗格苔蘚蟲、變口目、隱口目滅絕 | |
腕足動物門 | 96% | 正形貝目與長身貝目滅絕 | |
雙殼綱 | 59% | | |
腹足綱(蝸牛) | 98% | | |
菊石亞綱(頭足綱) | 97% | | |
海百合綱(棘皮動物門) | 98% | 游離海百合、圓頂海百合滅絕 | |
海蕾綱(棘皮動物門) | 100% | 可能在這次滅絕事件前不久滅絕 | |
三葉蟲綱 | 100% | 自泥盆紀開始衰退,在滅絕事件發生前僅有兩屬存活 | |
板足鱟綱(俗稱海蠍) | 100% | 可能在這次滅絕事件前不久滅絕 | |
介形綱(小型甲殼亞門) | 59% | | |
魚類 | |||
棘魚綱 | 100% | | |
海中生物
在二疊紀-三疊紀滅絕事件中,海生無脊椎動物的滅絕程度最大。在中國南部的一些富含化石的地層,在329屬的海生無脊椎動物中,有280屬在二疊紀最後兩個沉積層(以發現牙形石化石為標準)消失[1]。
根據統計分析的結果,二疊紀末海洋生物的物種數量降低,多是因為滅亡,而非物種形成的減少[38]。受到此次滅絕事件影響最嚴重的生物群,是具有碳酸鈣外殼的海洋無脊椎動物,尤其是吸收環境中二氧化碳以製造外殼的物種[39]。
在正常時期,底棲生物的滅亡率高。這次滅絕事件使底棲生物的滅亡率,達到正常時期的數倍[40][41]。以滅絕比例來看,海洋生物遭受災難性的滅亡[42][7][1][43]。
繼續存活的海生無脊椎動物則包含:腕足動物門(牠們的數量在滅絕事件後持續了一段緩慢的衰退)、菊石亞綱的齒菊石目與海百合綱(幾乎滅亡,但後來再度繁盛、多樣化)。
存活比例最高的海洋生物,通常具有良好的循環系統、複雜的氣體交換機制、以及较少的鈣化组织。而鈣化组织最多的物種,則是滅亡比例最高的生物[44][45]。以腕足動物為例,繼續存活的物種體型較小,且是多樣化生物群落中的數量稀少成員[46]。
自從二疊紀中期羅德階之後,菊石亞綱已經歷長時間的衰退,達約3000萬年之久。瓜達鹿白階(Guadalupian)末期的有限度滅絕事件,使菊石的差異性大幅減少,環境因素可能導致這次的滅絕事件。菊石的低多樣性與低差異性持續到二疊紀末滅絕事件;然而,二疊紀末滅絕事件的範圍廣泛、過程迅速。在三疊紀,菊石開始迅速多樣化,但物種之間的差異性仍然不大[47]。
在二疊紀時期,菊石的形態空間逐漸有限。在進入三疊紀的數百萬年後,菊石再度發展出原始的形態空間,但不同演化支已出現差異[48]。
陸地無脊椎動物
在二疊紀時期,陸地的昆蟲與其他無脊椎動物非常繁盛、多樣化,其中也包含地表上曾出現過的最大型昆蟲。發生在二疊紀末的滅絕事件,是目前已知惟一的昆蟲大規模滅絕事件[4],其中有八、九個昆蟲的目消失,而至少十個目的物種減少。古網翅總目(Palaeodictyopteroidea)在二疊紀中期開始衰退,這段時間可能與植物群的變化有關係。昆蟲的衰退高峰發生於二疊紀晚期,這段時間的高峰則可能與植物群的變化(由氣候變化造成)無直接關聯[7]。
在二疊紀-三疊紀滅絕事件的前後,昆蟲的化石有極大的差異。許多古生代的昆蟲多在二疊紀-三疊紀滅絕事件後消失,只有舌鞘目、Miomoptera(內翅總目的一目)、原直翅目繼續存活到中生代。外翅總目、單尾目、古網翅總目、Protelytroptera(舌鞘目的一亞目)、原蜻蜓目在二疊紀末滅亡。在一些保存狀態良好的三疊紀晚期地層中,通常可發現與現代昆蟲族群相關的化石[4]。
陸地植物
植物生態系統
陸地植物的化石紀錄很少見,大多數是花粉與孢子。所有大型滅絕事件對植物造成的影響,只有到科的層級[15]。若以種層級來看,減少的物種大部分可能是埋藏過程造成的假象[15]。若以優勢物種與地理分佈來看,可發現植物生態系統有重大的變動[15]。
在二疊紀/三疊紀交界時,優勢的植物群改變,許多陸地植物突然開始衰退,例如荷達樹(裸子植物)與舌羊齒(種子蕨)[49]。在二疊紀/三疊紀交界之後,原本佔優勢的裸子植物被石松門取代[50]。
一個針對格陵蘭東部沉積層中的花粉與孢子研究,顯示在滅絕事件發生前,當地有密集的裸子植物森林。在海生無脊椎動物群衰退時,這個大型森林也開始消失,而小型草本植物開始多樣化(包含石松門的卷柏與水韭)。裸子植物後來一度興盛,然後大量滅亡。在二疊紀末滅絕事件期間與之後,植物群發生數次興衰循環。優勢植物群從大型木本植物變動到草本植物,顯示當時許多的大型植物遭到環境改變的壓力。植物群落的接續與滅絕,發生於碳13/碳12比例極速下降之後,但兩者時間並不一致[51]。裸子植物森林的復原,花了約400萬到500萬年[15]。
煤層缺口
在三疊紀早期的地層中,沒有發現煤礦床,而三疊紀中期的地層中,煤礦床薄,而且品質低[52]。關於煤層缺口的成因有許多假設。有科學家認為,當時有許多新的真菌、昆蟲、以及脊椎動物出現,這些具侵略性的動物造成植物的大量死亡。但是,這些動物也因二疊紀-三疊紀滅絕事件而大量滅亡,不太可能是煤層缺口的主因[52]。有可能所有可形成煤礦的植物,在這次滅絕事件中幾乎滅亡;而在1000萬年之後,新的植物才適應潮濕、酸性的泥炭沼澤,而出現可形成煤礦的新植物[52]。非生物性因素,則包含:減少的降雨、或增加的碎屑沉積物[15]。最後,三疊紀早期的各種沉積層很少被發現,煤層的缺口可能反映了沉積層的稀少狀況。有可能形成煤礦的三疊紀早期生態系統,目前還沒有被發現[15]。例如在澳洲東部,當時可能長時間處在寒冷的氣候,這種天氣狀況適合泥炭的形成。在可以形成泥炭的植物中,有95%在這次滅絕事件中消失[53]。但是,澳洲與南極洲的煤礦床在P-Tr事件發生前,就已經消失[15]。
陸地脊椎動物
在二疊紀末期,許多陸地脊椎動物消失,而某些演化支甚至幾乎滅亡。某些存活的族系,並未長時間繼續存在,而其他族系則開始多樣化,並長期存活着。有充足證據顯示,超過⅔的陸生兩棲動物、蜥形綱、獸孔目的科在這個滅絕事件中消失。大型的草食性動物遭受嚴重的打擊。除了前稜蜥科,幾乎所有的二疊紀無孔亞綱動物滅亡;龜鱉目具有無孔類的頭骨,被認為可能是從前稜蜥科的近親演化而來(或是雙孔亞綱)。盤龍目早在二疊紀末期前就已滅亡。由於二疊紀的雙孔亞綱化石很少,無法確定滅絕事件對雙孔亞綱所造成的影響;雙孔亞綱包含大部分的爬行動物,例如蜥蜴、蛇、鱷魚、恐龍、以及恐龍的後代鳥類[54][55]。
滅絕模式的可能解釋
滅亡程度最嚴重的海生動物,包含:可以製造鈣質外殼(例如碳酸鈣)、代謝率低、以及呼吸系統較弱的物種,尤其是鈣質海綿類、四射珊瑚、床板珊瑚、具鈣質外殼的腕足動物、苔蘚動物門、棘皮動物門,上述動物中約有81%的屬滅亡。而無法製造鈣質外殼的近親,受到的影響較小,例如海葵(演化出現代的珊瑚)。具有高代謝率、呼吸系統良好、以及缺乏鈣質外殼的動物,滅亡的幅度較小;牙形石是個例外,約有33%的屬消失[56]。
這種滅絕模式,與組織缺氧產生的效應相符合。在當時,幾乎所有的大陸棚海域都有嚴重的缺氧現象,但缺氧現象無法解釋有限度的滅絕模式;因此缺氧不是造成海生生物滅絕的唯一成因。一個針對二疊紀晚期到三疊紀早期大氣層的數學模型,指出在這段期間,大氣層中的氧氣有明顯、長時期的衰退;這個衰退不因二疊紀/三疊紀交界而增加,並在三疊紀早期達到最低點,但仍高於現今的比例。由此得知,氧氣比例的衰退與滅絕模式並不一致[56]。
另一方面,海生生物的滅絕模式也與高碳酸血症(二氧化碳過高)產生的效應相符合。當二氧化碳濃度高於正常時,將對生物造成傷害:降低呼吸色素攜帶氧到組織的能力、使體液更為酸性、妨礙動物形成碳酸鈣外殼;當濃度更高時,會使生物進入昏迷狀態。除了上述反應,二氧化碳濃度高,會使生物製造鈣質外殼更加困難。與陸地生物相比,海洋生物對於二氧化碳濃度的變化相當敏感。溶解於水中的二氧化碳,是氧的28倍。對於進行呼吸作用的動物,二氧化碳的排除,與氣體經過呼吸系統膜相關;所以海洋動物會將身體內的二氧化碳保持在低濃度,低於陸地動物。若二氧化碳濃度增加,會妨礙海洋動物合成蛋白質、降低受精機率、形成不完整的鈣質外殼[56]。
目前很少發現跨越二疊紀與三疊紀交界的陸相地層,因此難以詳細研究陸地生物的滅絕/存活比例。二疊紀與三疊紀的昆蟲有顯著的不同,但是在二疊紀晚期到三疊紀早期,有個約1500萬年的昆蟲化石斷層。目前最足以研究二疊紀/三疊紀陸地脊椎動物變化的地層,位於南非的卡魯盆地(Karoo);但目前仍沒有足夠的研究可供結論[56]。
生態系統的復原
早期的研究認為在二疊紀末滅絕事件後,地表的生物迅速的復原,但這些研究多是針對在滅絕存活的先驅生物(Pioneer organisms)的研究結果,例如水龍獸。近年的大部分研究認為,當時經歷相當長的時間,生態系統才恢復以往的高度多樣性、複雜的食物鏈、與眾多生態位。在滅絕事件後,發生數次的滅絕波動,限制棲息地的復原,因此生態系統花了長時期才完全復原。近年的研究指出,地表生態系統直到進入三疊紀中期,約是滅絕事件後400萬到600萬年,才開始復原[57]。某些研究人員更指出,直到滅絕事件後3000萬年,進入三疊紀晚期,地表生態系統才完全復原[58]。

懷馬卡里里河,分布於紐西蘭南島,屬於辮狀河
在三疊紀早期,約是滅絕事件後400萬到600萬年,陸地的植物數量不足,造成煤層缺口,使當時的草食性動物面臨食物短缺[52]。南非卡魯盆地的河流,從曲折河改變為辮狀河,顯示該地區的植被曾長時間非常稀少[59]。
三疊紀早期的主要生態系統,無論是植物、動物,或是海洋、陸地系統,優勢生物都是少數特定、且全球分佈的物種。舉例而言,獸孔目的水龍獸,這種草食性動物的數量,佔了當時陸地草食性動物約90%;以及雙殼綱的克氏蛤(Claraia)、正海扇(Eumorphotis)、蚌形蛤(Unionites )、Promylina。正常的生態系統應由眾多物種構成,並佔據者不同的棲息地與生態位[49][60]。
在滅絕事件存活下來的某些劣勢物種,數量與棲息地大幅增加。例如:雙殼綱的海豆芽(Lingula)、出現於奧陶紀的疊層石、石松門的肋木(Pleuromeia)、種子蕨的二叉羊齒(Dicrodium)。[12][61][60][62]
海洋生態系統的改變

在二疊紀末滅絕事件後,濾食性固著動物(例如海百合)的數量大幅減少
在二疊紀-三疊紀滅絕事件發生前,接近67%的海生動物是固著動物,附著於海床生存;在中生代期間,只有約50%的海生動物是固著動物,其餘皆是可自由行動的動物。研究顯示,事件發生後,棲息在海底表層的濾食性固著動物,例如腕足動物、海百合,物種數量減少;而構造較複雜的動物,例如蝸牛、海膽、螃蟹,物種數量增加。
在二疊紀末滅絕事件發生前,同時存在複雜與單純的海洋生態系統,比例相當;生態系統經過復原後,複雜的海洋生物群落在數量上超過單純的海洋生物群落,比例相當於3/1[63];這使海生掠食動物面臨食物競爭的壓力,促成中生代海洋生物的進一步演化。
在滅絕事件發生前,雙殼綱是非常罕見的物種。在三疊紀,雙殼綱成為數量眾多、多樣化的生物群;其中的厚殼蛤(Rudists),是形成中生代時期礁岩的主要生物。部分研究人員認為雙殼綱的興起,發生於兩次滅絕高峰的間期[64]。
在滅絕事件中,海百合綱的物種數量、多樣性大幅減少,但沒有完全滅亡[65]。在滅絕事件後,海百合綱接着發生適應輻射,除了數量、棲息地增加,也發展出柔軟的腕足、具可動性[66]。
陸地脊椎動物
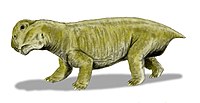
在三疊紀早期,水龍獸是陸地上最繁盛的脊椎動物。
水龍獸屬於獸孔目二齒獸類,是種體型接近豬的草食性動物,在三疊紀最早幾個時期,佔據了90%的陸地動物生態位[12]。除此之外,還有體型較小的肉食性犬齒獸類倖存,犬齒獸類是哺乳動物的祖先。根據非洲南部卡魯盆地的化石紀錄,有少數二疊紀物種也存活過二疊紀-三疊紀滅絕事件,例如獸頭亞目的四犬齒獸、犬齒獸 (Tetracynodon)、麝喙獸(Moschorhinus)、Ictidosuchoides,但數量沒有水龍獸繁盛[67]。
在三疊紀的早期,主龍類的數量遠少於獸孔目;但在三疊紀中期,主龍類取代獸孔目的優勢生態位[12]。在三疊紀中到晚期,主龍類的恐龍演化出現,並逐漸成為中生代的優勢陸地動物[68]。獸孔目與主龍類的興衰,與哺乳類的演化過程有關。獸孔目與隨後出現的哺乳形類,在這個時期多半演化成小型、夜行性的食蟲動物。夜間的生活習性,至少使哺乳形類發展出毛髮與較高的代謝率[69]。
兩棲動物離片椎目的某些物種,在滅絕事件中幾乎滅亡,但快速復原。在三疊紀的大部分時間,蝦蟆螈(Mastodonsaurus)與長吻迷齒螈下目(Trematosauria)是主要的水生與半水生掠食動物,以其他四足類、魚類為食[70]。
陸地脊椎動物經過相當長的時間,才從二疊紀末滅絕事件復原。一位研究人員估計陸地脊椎動物花了3000萬年,直到三疊紀晚期,才恢復之前的繁盛與多樣性。此時的陸地脊椎動物包含:兩棲動物、原始主龍類、恐龍、翼龍目、早期鱷魚、以及哺乳形類[3]。
可能的滅絕原因
關於二疊紀-三疊紀滅絕事件的發生過程,目前已有多種假設,包含劇烈與緩慢的過程;白堊紀-第三紀滅絕事件的發生過程,也有類似的假設。劇烈過程理論的成因,包含大型或多顆隕石造成的撞擊事件、連續性火山爆發、或是海床急驟釋放出大量甲烷水合物。緩慢過程理論的成因,包含海平面改變、缺氧、以及逐漸增加的乾旱氣候[12]。
陨石撞擊事件

位於南極洲的威爾克斯地隕石坑,直徑達500公里,形成時間在5億年內,規模與時代使它成為本次滅絕事件的可疑成因,但證據不夠明確。
白堊紀-第三紀滅絕事件由撞擊事件造成的證據,促使科學家們推論其他滅絕事件由撞擊事件造的的可能性,尤其是二疊紀-三疊紀滅絕事件。因此,科學家們盡力尋找那個時代的大型隕石坑與撞擊證據。
目前已在部分二疊紀/三疊紀交界的地層,發現撞擊事件的證據,例如:在澳洲與南極洲發現罕見的衝擊石英[71][72]、富勒烯包覆的外太空惰性氣體[73]、南極洲發現的玻璃隕石[74],以及地層中常見鐵、鎳、矽微粒[75]。但是,上述證據的真實性多受到懷疑[76][77][78][79]。在南極洲石墨峰(Graphite Peak)發現的衝擊石英,經過光學顯微鏡與穿透式電子顯微鏡重新檢驗後,發現其中的結構並非由撞擊產生,而是形變與地殼活動(例如火山)造成的[80]。
目前已經發現數個可能與二疊紀末滅絕事件有關的隕石坑,包含:澳洲西北外海的貝德奧高地[72]、南極洲東部的威爾克斯地隕石坑[81][82]。但沒有可信服的證據,可證明這兩個地形是由撞擊產生。以威爾克斯地隕石坑為例,這個位在冰原下的凹地,年代無法確定,可能晚於二疊紀末滅絕事件才形成。其中最爭議的隕石坑,是由地質學家Michael Stanton在2002年提出的,他主張墨西哥灣是形成於二疊紀末期的一次撞擊事件,而該撞擊事件也造成了二疊紀-三疊紀滅絕事件[83]。
如果二疊紀末滅絕事件的主因是由於撞擊,很有可能隕石坑已經消失於地表。地球表面有70%是海洋,所以隕石或彗星撞擊海洋的機率,是撞擊陸地的兩倍以上。但是,地球的海洋地殼會因聚合與隱沒作用而消失於地表,所以目前無法找到距今2億年以上的海洋地殼。如果當時有非常大型的撞擊事件,撞擊會使該處地殼破裂、變薄,造成大量的熔岩[84]。
撞擊理論最受到關注的原因是,它可與其他滅絕現象產生因果連結[85],例如西伯利亞暗色岩火山爆發,可能由大型撞擊產生[86],甚至是大型撞擊的對蹠點[87][85]。即使撞擊發生於海洋,隕石坑因為隱沒帶而消失於地表,應會留下其他證據。如同白堊紀-第三紀滅絕事件,如果有撞擊事件發生,會產生大量撞擊拋出物,相同時代的許多地層會發現大量親鐵元素,例如銥。若滅絕事件由隕石撞擊引發,可以解釋在滅絕後,生物沒有快速的適應演化。
火山爆發

西伯利亞暗色岩的熔岩範圍
在二疊紀的最後一期,發生兩個大規模火山爆發:西伯利亞暗色岩、峨嵋山暗色岩。峨嵋山暗色岩位於現今中國四川省,規模較小,形成時間是瓜達鹿白階末期,形成時的位置接近赤道[88][89]。西伯利亞暗色岩火山爆發是地質史上已知最大規模的火山爆發之一,熔岩面積超過200萬平方公里。西伯利亞暗色岩火山爆發原本被認為持續約數百萬年,但近年的研究認為西伯利亞暗色岩形成於2億5120萬年前(誤差值為30萬年),接近二疊紀末期[1][90]。
峨嵋山暗色岩與西伯利亞暗色岩火山爆發,可能製造大量灰塵與酸性微粒,遮蔽照射到地表的陽光,妨礙陸地與海洋透光帶的生物進行光合作用,進而遭成食物鏈的崩潰。大氣層中的酸性微粒,最後形成酸雨降落到地表。酸雨對陸地植物、可製造碳酸鈣硬殼的軟體動物與浮游生物造成傷害。火山爆發也釋放大量二氧化碳,形成溫室效應。大氣層中的灰塵與酸性物質降落到地表之後,過量的二氧化碳持續形成溫室效應[85]。
與其他火山相比,西伯利亞暗色岩更為危險。洪流玄武岩會產生大量的流動性熔岩,只會噴發少量的蒸氣、碎屑進入大氣層。但是,西伯利亞暗色岩火山爆發的噴出物質,似乎有20%是火山碎屑,這些火山灰與火山礫進入大氣層後,會造成短時期的氣候寒冷[91]。帶有玄武岩的熔岩侵入碳酸鹽岩、或帶有大型煤層的地區,會產生大量的二氧化碳,會在大氣層的灰塵降落到地表後,造成全球暖化[85]。
這些火山爆發事件的規模是否足以造成二疊紀末滅絕事件,仍有爭議。峨嵋山暗色岩接近赤道區,火山爆發所製造的灰塵與酸性物質,會對全世界造成影響。西伯利亞暗色岩的規模較大,但位置在北極區內,或在北極區附近。如果西伯利亞暗色岩火山爆發的持續時間在20萬年內,會使大氣層中的二氧化碳達到正常程度的兩倍。近年的氣候模型顯示,大氣層中的二氧化碳含量加倍,會使全球氣候上升1.5°C到4.5°C,這會造成嚴重的影響,但沒有到二疊紀-三疊紀滅絕事件的嚴重程度[85]。
在2005年,日本NHK與加拿大國家電影委員會(NFB)製作的《地球大進化》(Miracle Planet;地球大進化〜46億年・人類への旅)電視節目,提出火山爆發形成的輕微全球暖化,導致甲烷水合物的氣化;由於甲烷氣體對全球暖化的影響,是二氧化碳的45倍,甲烷水合物的氣化進而導致不斷循環的全球暖化。
甲烷水合物的氣化
科學家們已在全球許多地點的二疊紀末碳酸鹽礦層中,發現當時的碳13/碳12比例有迅速減少的跡象,減少了約10‰左右[92][43]。從二疊紀末開始,碳13/碳12比例發生了一系列的上升與下降現象,直到三疊紀中期才穩定、停止;而發生於二疊紀末的第一次變動,是其中規模最大、最迅速的變動。在三疊紀中期,碳13/碳12比例穩定之後,可製造碳酸鈣外殼的動物開始復原[44]。
碳13/碳12比例的下降,可能有以下多種因素[93]:
- 火山爆發產生的氣體,其碳13/碳12比例低於正常值約5~8‰。但若要使全球的碳13/碳12比例下降約10‰,其火山爆發的規模將超越目前已知的任何地質紀錄[94]。
- 生物活動降低,使環境中的碳12更慢被攝取,更多碳12進入沉積層,而使碳13/碳12比例降低。所有的化學反應是建立在原子間的電磁力。較輕的同位素,其化學反應較快。所以生物化學過程會用到較輕的同位素。但關於古新世-始新世交替時期最大熱量(PETM)事件的研究發現,該時期的碳13/碳12比例小幅降低約3到4‰。根據假設,即使將全部的有機碳(包含生物、土壤、海洋)進入土壤沉積層中,也不會達到古新世/始新世交替時的小幅度碳13/碳12比例下降。由此可知生物活動降低不是二疊紀末期的碳13/碳12比例降低的原因[94]。
- 埋在沉積層中的死亡生物,體內的碳13/碳12比例小於正常值約20到25‰。就理論上而言,如果海平面迅速降低,淺海地區的沉積層曝露到空氣後,開始氧化作用。但若要使全球的碳13/碳12比例下降約10‰,要有6.5到8.4兆噸有機碳經氧化後形成,而沉積層本身需要數十萬年的氧化。這似乎不太可能發生[7]。
- 間歇性海洋高氧與缺氧事件,也可能是三疊紀早期的碳13/碳12比例下降的原因[44]。全球性的海洋缺氧現象,本身也是滅絕事件的原因之一。二疊紀末期到三疊紀早期的陸地,多為熱帶地區。熱帶的大型河流會將沉積層中有有機碳帶入海洋,尤其是低緯度的鄰近海盆。生物化學過程會用到較輕的同位素,所以有機碳的碳13/碳12比例低。大量有機碳的迅速(以相對而言)釋放與沉降,可能會引發間歇性的高氧/缺氧事件[95]。元古宙晚期到寒武紀交接時期,也曾發生碳13/碳12比例的下降,可能與此相關,或是其他與海洋相關的因素[44]。

甲烷水合物即俗稱的可燃冰,大部分位在大陸棚,被視為未來的潛在能源之一。
其他的理論則有:海洋大量釋放二氧化碳[96]、以及全球的碳循環系統經歷長時間的重整[93]。
但最有可能導致全球性碳13/碳12比例下降的因素,是甲烷水合物氣化產生的甲烷[7]。而利用碳循環模型模擬的結果,甲烷最有可能導致如此大幅的下降[93][96]。甲烷水合物是固態形式的水於晶格(水合物)中包含大量的甲烷。甲烷是由甲烷菌製造,碳13/碳12比例低於正常值約60‰。甲烷在特定的壓力與溫度下,會形成包合物,例如永凍層的近表層,並在大陸棚、更深的海床等地區大量形成。甲烷水合物通常出現在海平面300公尺以下的沉積層。最深可在水深2000公尺處發現,但大部分在水深1100公尺以上[97]。
西伯利亞暗色岩火山爆發產生的熔岩面積,是以往認定的兩倍以上,新發現的熔岩地區,在二疊紀末時幾乎是淺海。極有可能這些淺海地區蘊含甲烷水合物,而火山爆發產生的熔岩流入海床後,促使甲烷水合物的汽化[98]。
由於甲烷本身是種非常強的溫室氣體,大量的甲烷被視為造成全球暖化的主要原因。證據顯示該時期的全球氣溫上升,赤道區上升約6°C,高緯度地區上升更多。例如:氧18/氧16比例的下降[99]、舌羊齒植物群(舌羊齒與生存於相同地區的植物)消失,由生存於低緯度的植物群取代[100][12]。
碳13/碳12比例的變動,被認為與大量釋放的甲烷有關,但兩者在三疊紀早期的變動模式,並不吻合。要造成如此大的氣候變遷,所需要的甲烷量,是引發古新世-始新世交替時期最大熱量的五倍[44]。但三疊紀早期曾出現大量碳13,使碳13/碳12比例迅速增加,而後下降[44]。
海平面改變
當原本沉浸的海床露出海平面時,會造成海退。海平面的下降會使淺海的生存區域減少,破壞當地的生態系統。淺海的可棲息地,富含食物鍊下層的生物,這些生物的減少,使賴其以維生的生物競爭食物更激烈[101]。海退與滅絕事件之間似乎有部份關聯,但另有證據認為兩者間沒有關係,而海退會形成新的棲息地。[12]海平面的變化,同時也導致海底沉積物的變化,並影響海水溫度與鹽度,進一步造成海生生物的多樣性衰退[102]。
海洋缺氧
有證據顯示,二疊紀末期的海洋發生了缺氧事件。在格陵蘭東部的一個二疊紀末期海相沉積層,指出當時有明顯、快速的海洋缺氧現象[103]。而數個二疊紀末沉積層的鈾/釷比例,也指出在這次滅絕事件發生時,海洋有嚴重的缺氧現象[104]。
缺氧事件可能導致海洋生物的大量死亡,只有棲息於海底泥層、可以進行無氧呼吸的細菌不受影響。另有證據顯示,這次海洋缺氧事件,造成海床大量釋放硫化氫。
海洋缺氧事件的原因,可能是長時間的全球暖化,降低赤道區與極區之間的溫度梯度,進而造成溫鹽環流系統的緩慢,甚至停止。溫鹽環流系統的緩慢或停止,可能使得海洋中的含氧量減少[104]。
但是,某些研究人員架構出二疊紀末期的海洋溫鹽環流系統,認為當時的溫鹽環流系統無法解釋深海區域的缺氧現象[105]。
硫化氫
二疊紀末期發生的海洋缺氧事件,可能使硫酸鹽還原菌成為海洋生態系統中的優勢物種,包含脫硫桿菌目、脫硫弧菌目、互營桿菌目、熱脫硫桿菌門,這些生物會製造大量的硫化氫,過量的硫化氫會對陸地、海洋中的動植物造成毒害,並破壞臭氧層,使生物暴露在紫外線下[106]。在二疊紀末到三疊紀早期發現許多綠菌,它們進行不產氧光合作用,釋放出硫化氫。綠菌的興盛時期,與二疊紀末滅絕事件和事後的長期復原,時期相符[107]。大氣層中的二氧化碳增加,植物卻大規模滅亡,硫化氫理論可以解釋植物的大規模滅亡。二疊紀末地層中的孢子化石,多數帶有不正常特徵,可能是由硫化氫破壞臭氧層,大量的紫外線進入地表造成。
盤古大陸的地圖
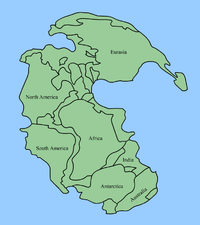
盤古大陸的形成
在二疊紀中期(約是烏拉爾世的空谷階),幾乎地表的所有大陸聚合成盤古大陸,盤古大陸由泛大洋環繞着,而東亞部份直到二疊紀末期才與盤古大陸聚合[108]。盤古大陸的形成,使得全球大部分的淺水區域消失,而淺水區域是海洋中最多生物棲息的部份。原本隔離的大陸架連接之後,使彼此獨立的生態系統開始互相競爭。盤古大陸的形成,造成了單一的海洋循環系統,以及單一的大氣氣候系統,在盤古大陸的海岸形成季風氣候,而廣大的內陸則形成乾旱的氣候。
在盤古大陸形成後,海洋生物數量減少,滅亡比例接近其他大型滅絕事件。盤古大陸的形成,似乎對陸地生物沒有造成嚴重的變化,二疊紀晚期的獸孔目反而因此擴大生存領域、更為多樣性。盤古大陸的形成,使海洋生物開始減少,但不是導致二疊紀末滅絕事件的直接原因。
多重原因
二疊紀-三疊紀末滅絕事件的產生原因,可能由上述事件連鎖、交錯形成,並日趨嚴重。西伯利亞暗色岩的火山爆發,除了產生大量的二氧化碳與甲烷,也破壞鄰近地區的煤層與大陸架[56]。接下來的全球暖化,間接導致地質歷史中最嚴重的海洋缺氧事件。海洋的缺氧,使硫化菌等進行不產氧的化合作用生物興起,它們釋放出大量的硫化氫[56]。
但是,這連鎖、交錯的事件,部分環節相當薄弱。碳13/碳12比例的變動,被認為與大量釋放的甲烷有關,但兩者在三疊紀早期的變動模式,並不吻合[44]。二疊紀末期的海洋溫鹽環流系統,不會造成深海區域的缺氧事件[105]。
参考文献
^ 1.01.11.21.31.41.51.6 (英文)
Jin YG, Wang Y, Wang W, Shang QH, Cao CQ, Erwin DH. Pattern of Marine Mass Extinction Near the Permian–Triassic Boundary in South China. Science. 2000, 289 (5478): 432–436. PMID 10903200. doi:10.1126/science.289.5478.432.
^ 2.02.1 (英文)Bowring SA, Erwin DH, Jin YG, Martin MW, Davidek K, Wang W. U/Pb Zircon Geochronology and Tempo of the End-Permian Mass Extinction. Science. 1998, 280 (1039): 1039–1045. doi:10.1126/science.280.5366.1039.
^ 3.03.13.2 (英文)Benton M J. When Life Nearly Died: The Greatest Mass Extinction of All Time. Thames & Hudson. 2005. ISBN 978-0500285732.
^ 4.04.14.2 (英文)Labandeira CC, Sepkoski JJ. Insect diversity in the fossil record. Science. 1993, 261 (5119): 310–5. PMID 11536548. 使用|accessdate=需要含有|url=(帮助)
^ (英文)Sole, R. V., and Newman, M., 2002. "Extinctions and Biodiversity in the Fossil Record - Volume Two, The earth system: biological and ecological dimensions of global environment change" pp. 297-391, Encyclopedia of Global Enviromental Change John Wilely & Sons.
^ (英文)Barry, Patrick L. The Great Dying. Science@NASA. Science and Technology Directorate, Marshall Space Flight Center, NASA. January 28, 2002 [March 26, 2009]. (原始内容存档于2012年2月16日).
^ 7.07.17.27.37.4 (英文)Erwin DH. The great Paleozoic crisis; Life and death in the Permian. Columbia University Press. 1993. ISBN 0231074670.
^ (英文)Yin H, Zhang K, Tong J, Yang Z, Wu S. The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary. Episodes: 102–114.
^ (英文)Yin HF, Sweets WC, Yang ZY, Dickins JM,. Permo-Triassic Events in the Eastern Tethys. Cambridge Univ. Pres, Cambridge, 1992.
^ 超級火山噴發造成史前地球生物滅絕
^
科學家找到地球生命大滅絕證據. [2011-01-25]. (原始内容存档于2011-01-31). 互联网档案馆的存檔,存档日期2011-01-31.
^ 12.012.112.212.312.412.512.6 (英文)Tanner LH, Lucas SG & Chapman MG. Assessing the record and causes of Late Triassic extinctions (PDF). Earth-Science Reviews. 2004, 65 (1-2): 103–139 [2007-10-22]. doi:10.1016/S0012-8252(03)00082-5. (原始内容 (PDF)存档于2007-10-25).
^ Burgess, S. D.; Bowring, S.; Shen, S.-z. High-precision timeline for Earth's most severe extinction. Proceedings of the National Academy of Sciences. 2014-02-10, 111 (9): 3316–3321. doi:10.1073/pnas.1317692111. 引文使用过时参数coauthors (帮助)
^ (英文)Erwin, D.H. The Great Paleozoic Crisis: Life and Death in the Permian. New York isbn=0231074670: Columbia University Press. 1993.
^ 15.015.115.215.315.415.515.615.715.8 (英文)McElwain, J.C.; Punyasena, S.W. Mass extinction events and the plant fossil record. Trends in Ecology & Evolution. 2007, 22 (10): 548–557. doi:10.1016/j.tree.2007.09.003. 引文使用过时参数coauthors (帮助)
^ (英文)Magaritz, M. 13C minima follow extinction events: a clue to faunal radiation. Geology. 1989, 17: 337–340.
^ (英文)Krull, S.J., and Retallack, J.R. 13C depth profiles from paleosols across the Permian–Triassic boundary: Evidence for methane release. GSA Bulletin. 2000, 112 (9): 1459–1472. ISSN 0016-7606. doi:10.1130/0016-7606(2000)112<1459:CDPFPA>2.0.CO;2.
^ (英文)Dolenec, T., Lojen, S., Ramovs, A. The Permian–Triassic boundary in Western Slovenia (Idrijca Valley section): magnetostratigraphy, stable isotopes, and elemental variations. Chemical Geology. 2001, 175 (1): 175–190. doi:10.1016/S0009-2541(00)00368-5.
^ (英文)Musashi, M., Isozaki, Y., Koike, T. and Kreulen, R. Stable carbon isotope signature in mid-Panthalassa shallow-water carbonates across the Permo–Triassic boundary: evidence for 13C-depleted ocean. Earth Planet. Sci. Lett. 2001, 193: 9–20. doi:10.1016/S0012-821X(01)00398-3.
^ (英文)Dolenec, T., Lojen, S., and Ramovs, A. The Permian-Triassic boundary in Western Slovenia (Idrijca Valley section): magnetostratigraphy, stable isotopes, and elemental variations. Chemical Geology. 2001, 175: 175–190. doi:10.1016/S0009-2541(00)00368-5.
^ 21.021.1 (英文)H Visscher, H Brinkhuis, D L Dilcher, W C Elsik, Y Eshet, C V Looy, M R Rampino, and A Traverse. The terminal Paleozoic fungal event: Evidence of terrestrial ecosystem destabilization and collapse. Proceedings of the National Academy of Sciences. 1996, 93 (5): 2155–2158 [2007-09-20].
^ (英文)Foster, C.B.; Stephenson, M.H.; Marshall, C.; Logan, G.A.; Greenwood, P.F. A Revision Of Reduviasporonites Wilson 1962: Description, Illustration, Comparison And Biological Affinities. Palynology. 2002, 26 (1): 35–58. doi:10.2113/0260035. 引文使用过时参数coauthors (帮助)
^ (英文)López-Gómez, J. and Taylor, E.L. Permian-Triassic Transition in Spain: A multidisciplinary approach. Palaeogeography, Palaeoclimatology, Palaeoecology. 2005, 229 (1-2): 1–2. doi:10.1016/j.palaeo.2005.06.028. (原始内容存档于2010-03-06).
^ (英文)Looy, C.V.; Twitchett, R.J.; Dilcher, D.L.; Van Konijnenburg-van Cittert, J.H.A.; Visscher, H. Life in the end-Permian dead zone. Proceedings of the National Academy of Sciences. 2005, 162 (4): 653–659. PMID 11427710. doi:10.1073/pnas.131218098.See image 2
引文使用过时参数coauthors (帮助)
^ 25.025.1 (英文)Ward PD, Botha J, Buick R, De Kock MO, Erwin DH, Garrison GH, Kirschvink JL & Smith R. Abrupt and Gradual Extinction Among Late Permian Land Vertebrates in the Karoo Basin, South Africa. Science. 2005, 307 (5710): 709–714. doi:10.1126/science.1107068.
^ (英文)Retallack, G.J.; Smith, R.M.H.; Ward, P.D. Vertebrate extinction across Permian-Triassic boundary in Karoo Basin, South Africa. Bulletin of the Geological Society of America. 2003, 115 (9): 1133–1152. doi:10.1130/B25215.1. (原始内容存档于2008-05-05). 引文使用过时参数coauthors (帮助)
^ Sephton, Mark A.; Visscher, Henk; Looy, Cindy V.; Verchovsky, Alexander B.; Watson, Jonathon S. Chemical constitution of a Permian-Triassic disaster species. Geology. 2009, 37 (10): 875–878. doi:10.1130/G30096A.1. 引文使用过时参数coauthors (帮助)
^ (英文)Rampino MR, Prokoph A & Adler A. Tempo of the end-Permian event: High-resolution cyclostratigraphy at the Permian–Triassic boundary. Geology. 2000, 28 (7): 643–646. Bibcode:2000Geo....28..643R. ISSN 0091-7613. doi:10.1130/0091-7613(2000)28<643:TOTEEH>2.0.CO;2.
^ (英文)Wang, S.C.; Everson, P.J. Confidence intervals for pulsed mass extinction events. Paleobiology. 2007, 33 (2): 324–336. doi:10.1666/06056.1. 引文使用过时参数coauthors (帮助)
^ (英文)Twitchett RJ Looy CV Morante R Visscher H & Wignall PB. Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis. Geology. 2001, 29 (4): 351–354. doi:10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2. 引文格式1维护:日期与年 (link)
^ Retallack, G.J.; Metzger, C.A.; Greaver, T.; Jahren, A.H.; Smith, R.M.H.; Sheldon, N.D. Middle-Late Permian mass extinction on land. Bulletin of the Geological Society of America. 2006, 118 (11-12): 1398–1411. doi:10.1130/B26011.1. 引文使用过时参数coauthors (帮助)
^ (英文)Stanley SM & Yang X. A Double Mass Extinction at the End of the Paleozoic Era. Science. 1994, 266 (5189): 1340–1344. PMID 17772839. doi:10.1126/science.266.5189.1340.
^ (英文)Shen, S., and Shi, G.R. Paleobiogeographical extinction patterns of Permian brachiopods in the Asian-western Pacific region. Paleobiology. 2002, 28: 449–463. doi:10.1666/0094-8373(2002)028<0449:PEPOPB>2.0.CO;2.
^ (英文)Wang, X-D, and Sugiyama, T. Diversity and extinction patterns of Permian coral faunas of China. Lethaia. December 2000, 33 (4): 285–294. doi:10.1080/002411600750053853.
^ Retallack, G.J., Metzger, C.A., Jahren, A.H., Greaver, T., Smith, R.M.H., and Sheldon, N.D. Middle-Late Permian mass extinction on land. GSA Bulletin. November/December 2006, 118 (11/12): 1398–1411. doi:10.1130/B26011.1. 请检查|date=中的日期值 (帮助)
^ (英文)Ota, A, and Isozaki, Y. Fusuline biotic turnover across the Guadalupian–Lopingian (Middle–Upper Permian) boundary in mid-oceanic carbonate buildups: Biostratigraphy of accreted limestone in Japan. Journal of Asian Earth Sciences. March 2006, 26 (3-4): 353–368.
^ (英文)
Racki G. Silica-secreting biota and mass extinctions: survival processes and patterns. Palaeogeography Palaeoclimatology Palaeoecology. 1999, 154 (1–2): 107–132. doi:10.1016/S0031-0182(99)00089-9.
^ Bambach, R.K.; Knoll, A.H.; Wang, S.C., Origination, extinction, and mass depletions of marine diversity, Paleobiology, December 2004, 30 (4): 522–542
^ Knoll, A.H., Biomineralization and evolutionary history. In: P.M. Dove, J.J. DeYoreo and S. Weiner (Eds), Reviews in Mineralogy and Geochemistry, (PDF), (2004), (原始内容 (PDF)存档于2010-06-20) 请检查|date=中的日期值 (帮助)
^ (英文)Stanley, S.M. Predation defeats competition on the seafloor. Paleobiology. 2008, 34 (1): 1–21 [2008-05-13]. doi:10.1666/07026.1.
^ (英文)Stanley, S.M. An Analysis of the History of Marine Animal Diversity. Paleobiology. 2007, 33 (sp6): 1–55. doi:10.1666/06020.1.
^ (英文)McKinney, M.L. Taxonomic selectivity and continuous variation in mass and background extinctions of marine taxa. Nature. 1987, 325 (6100): 143–145. doi:10.1038/325143a0.
^ 43.043.1 (英文)Twitchett RJ, Looy CV, Morante R, Visscher H, Wignall PB. Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis. Geology. 2001, 29 (4): 351–354. doi:10.1130/0091-7613(2001)029%3C0351:RASCOM%3E2.0.CO;2.
^ 44.044.144.244.344.444.544.6 (英文)Payne, J.L.; Lehrmann, D.J.; Wei, J.; Orchard, M.J.; Schrag, D.P.; Knoll, A.H. Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction. Science. 2004, 305 (5683): 506. PMID 15273391. doi:10.1126/science.1097023. 引文使用过时参数coauthors (帮助)
^ (英文)Knoll, A.H.; Bambach, R.K.; Canfield, D.E.; Grotzinger, J.P. Comparative Earth history and Late Permian mass extinction. Science (Washington). 1996, 273 (5274): 452–456. PMID 8662528. doi:10.1126/science.273.5274.452. 引文使用过时参数coauthors (帮助)
^ (英文)Leighton, L.R.; Schneider, C.L. Taxon characteristics that promote survivorship through the Permian–Triassic interval: transition from the Paleozoic to the Mesozoic brachiopod fauna. Paleobiology. 2008, 34 (1): 65–79. doi:10.1666/06082.1. 引文使用过时参数coauthors (帮助)
^ (英文)Villier, L. Morphological Disparity of Ammonoids and the Mark of Permian Mass Extinctions. Science. 2004, 306: 264. doi:10.1126/science.1102127.
^ (英文)Saunders, W. B. Morphologic and taxonomic history of Paleozoic ammonoids in time and morphospace. Paleobiology. 2008, 34: 128. doi:10.1666/07053.1.
^ 49.049.1 (英文)Retallack, GJ. Permian–Triassic life crisis on land. Science. 1995, 267 (5194): 77–80. doi:10.1126/science.267.5194.77.
^ (英文)Looy, CV Brugman WA Dilcher DL & Visscher H. The delayed resurgence of equatorial forests after the Permian–Triassic ecologic crisis. Proceedings National Academy of Sciences. 1999, 96: 13857–13862. PMID 10570163.
^ (英文)Looy, CV; Twitchett RJ, Dilcher DL, &Van Konijnenburg-Van Cittert JHA and Henk Visscher. Life in the end-Permian dead zone. Proceedings of the National Academy of Sciences. July 3, 2001, 14 (98): 7879–7883. doi:10.1073/pnas.131218098. 引文使用过时参数coauthors (帮助)
^ 52.052.152.252.3 (英文)Retallack GJ Veevers JJ & Morante R. Global coal gap between Permian–Triassic extinctions and middle Triassic recovery of peat forming plants. GSA Bulletin. 1996, 108 (2): 195–207 [2007-09-29].
^ (英文)Michaelsen P. Mass extinction of peat-forming plants and the effect on fluvial styles across the Permian–Triassic boundary, northern Bowen Basin, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology. 2002, 179 (3–4): 173–188. doi:10.1016/S0031-0182(01)00413-8.
^ (英文)Maxwell, W. D. "Permian and Early Triassic extinction of non-marine tetrapods". Palaeontology. 1992, 35: 571–583.
^ Erwin DH. The End-Permian Mass Extinction. Annual Review of Ecology and Systematics. 1990, 21: 69–91. doi:10.1146/annurev.es.21.110190.000441.
^ 56.056.156.256.356.456.5 (英文)Knoll, A.H., Bambach, R.K., Payne, J.L., Pruss, S., and Fischer, W.W. Paleophysiology and end-Permian mass extinction. Earth and Planetary Science Letters. 2007, 256: 295–313 [2008-07-04]. doi:10.1016/j.epsl.2007.02.018. (原始内容存档于2008-04-10). Full contents may be available online at Paleophysiology and end-Permian mass extinction (PDF). [2008-07-04].
^ (英文)Lehrmann, D.J., Ramezan, J., Bowring, S.A.; 等. Timing of recovery from the end-Permian extinction: Geochronologic and biostratigraphic constraints from south China. Geology. December 2006, 34 (12): 1053–1056. doi:10.1130/G22827A.1. 引文格式1维护:显式使用等标签 (link)
^ (英文)Sahney, S. and Benton, M.J. Recovery from the most profound mass extinction of all time (PDF). Proceedings of the Royal Society: Biological. 2008, 275: 759. doi:10.1098/rspb.2007.1370.
^ Ward PD, Montgomery DR, & Smith R. Altered river morphology in South Africa related to the Permian–Triassic extinction. Science. 2000, 289 (5485): 1740–1743. doi:10.1126/science.289.5485.1740.
^ 60.060.1 (英文)Hallam A & Wignall PB. Mass Extinctions and their Aftermath. Oxford University Press. 1997. ISBN 978-0198549161.
^ (英文)Zi-qiang W. Recovery of vegetation from the terminal Permian mass extinction in North China. Review of Palaeobotany and Palynology. 1996, 91 (1–4): 121–142. doi:10.1016/0034-6667(95)00069-0.
^ Rodland, DL & Bottjer, DJ. Biotic Recovery from the End-Permian Mass Extinction: Behavior of the Inarticulate Brachiopod Lingula as a Disaster Taxon. Palaios. 2001, 16 (1): 95–101. doi:10.1669/0883-1351(2001)0163%C0095:BRFTEP3%E2.0.CO;2.
^ (英文)Wagner PJ, Kosnik MA, & Lidgard S. Abundance Distributions Imply Elevated Complexity of Post-Paleozoic Marine Ecosystems. Science. 2006, 314 (5803): 1289–1292. doi:10.1126/science.1133795.
^ (英文)Clapham, M.E., Bottjer, D.J. and Shen, S. Decoupled diversity and ecology during the end-Guadalupian extinction (late Permian). Geological Society of America Abstracts with Programs. 2006, 38 (7): 117 [2008-03-28].
^ (英文)Foote, M. Morphological diversity in the evolutionary radiation of Paleozoic and post-Paleozoic crinoids (PDF). Paleobiology. 1999, 25 (sp1): 1–116 [2008-05-12]. doi:10.1666/0094-8373(1999)25[1:MDITER]2.0.CO;2.
^ (英文)Baumiller, T. K. Crinoid Ecological Morphology. Annual Review of Earth and Planetary Sciences (PDF)使用|format=需要含有|url=(帮助). 2008, 36: 221–249. doi:10.1146/annurev.earth.36.031207.124116.
^
Botha, J., and Smith, R.M.H. Lystrosaurus species composition across the Permo–Triassic boundary in the Karoo Basin of South Africa. Lethaia. 2007, 40: 125–137 [2008-07-02]. doi:10.1111/j.1502-3931.2007.00011.x. [永久失效連結] Full version online at Lystrosaurus species composition across the Permo–Triassic boundary in the Karoo Basin of South Africa (PDF). [2008-07-02].
^ (英文)Benton, M.J. Vertebrate Paleontology. Blackwell Publishers. 2004: xii–452. ISBN 0-632-05614-2.
^ (英文)Ruben, J.A., and Jones, T.D. Selective Factors Associated with the Origin of Fur and Feathers. American Zoologist. 2000, 40 (4): 585–596. doi:10.1093/icb/40.4.585.
^ (英文)Yates AM & Warren AA. The phylogeny of the 'higher' temnospondyls (Vertebrata: Choanata) and its implications for the monophyly and origins of the Stereospondyli. Zoological Journal of the Linnean Society. 2000, 128 (1): 77–121 [2008-01-18]. (原始内容存档于2007-10-01).
^ (英文)Retallack GJ, Seyedolali A, Krull ES, Holser WT, Ambers CP, Kyte FT. Search for evidence of impact at the Permian–Triassic boundary in Antarctica and Australia. Geology. 1998, 26 (11): 979–982.
^ 72.072.1 (英文)Becker L, Poreda RJ, Basu AR, Pope KO, Harrison TM, Nicholson C, Iasky R. Bedout: a possible end-Permian impact crater offshore of northwestern Australia. Science. 2004, 304 (5676): 1469–1476. doi:10.1126/science.1093925.
^ (英文)Becker L, Poreda RJ, Hunt AG, Bunch TE, Rampino M. Impact event at the Permian–Triassic boundary: Evidence from extraterrestrial noble gases in fullerenes. Science. 2001, 291 (5508): 1530–1533. doi:10.1126/science.1057243.
^ (英文)Basu AR, Petaev MI, Poreda RJ, Jacobsen SB, Becker L. Chondritic meteorite fragments associated with the Permian–Triassic boundary in Antarctica. Science. 2003, 302 (5649): 1388–1392. doi:10.1126/science.1090852.
^ (英文)Kaiho K, Kajiwara Y, Nakano T, Miura Y, Kawahata H, Tazaki K, Ueshima M, Chen Z, Shi GR. End-Permian catastrophe by a bolide impact: Evidence of a gigantic release of sulfur from the mantle. Geology. 2001, 29 (9): 815–818 [2007-10-22].
^ (英文)Farley KA, Mukhopadhyay S, Isozaki Y, Becker L, Poreda RJ. An extraterrestrial impact at the Permian–Triassic boundary?. Science. 2001, 293 (5539): 2343. doi:10.1126/science.293.5539.2343a.
^ (英文)Koeberl C, Gilmour I, Reimold WU, Philippe Claeys P, Ivanov B. End-Permian catastrophe by bolide impact: Evidence of a gigantic release of sulfur from the mantle: Comment and Reply. Geology. 2002, 30 (9): 855–856. doi:10.1130/0091-7613(2002)030%3C0855:EPCBBI%3E2.0.CO;2.
^ (英文)Isbell JL, Askin RA, Retallack GR. Search for evidence of impact at the Permian–Triassic boundary in Antarctica and Australia; discussion and reply. Geology. 1999, 27 (9): 859–860. doi:10.1130/0091-7613(1999)027%3C0859:SFEOIA%3E2.3.CO;2.
^ (英文)Koeberl K, Farley KA, Peucker-Ehrenbrink B, Sephton MA. Geochemistry of the end-Permian extinction event in Austria and Italy: No evidence for an extraterrestrial component. Geology. 2004, 32 (12): 1053–1056. doi:10.1130/G20907.1.
^ (英文)Langenhorst F, Kyte FT & Retallack GJ. Reexamination of quartz grains from the Permian–Triassic boundary section at Graphite Peak, Antarctica (PDF). Lunar and Planetary Science Conference XXXVI. 2005 [2007-07-13].
^ (英文)von Frese RR, Potts L, Gaya-Pique L, Golynsky AV, Hernandez O, Kim J, Kim H & Hwang J. Permian–Triassic mascon in Antarctica. Eos Trans. AGU, Jt. Assem. Suppl. 2006, 87 (36): Abstract T41A–08 [2007-10-22]. (原始内容存档于2007-09-30).
^ (英文)Von Frese, R.R.B.; L. V. Potts, S. B. Wells, T. E. Leftwich, H. R. Kim, J. W. Kim, A. V. Golynsky, O. Hernandez, and L. R. Gaya-Piqué. GRACE gravity evidence for an impact basin in Wilkes Land, Antarctica. Geochem. Geophys. Geosyst. 2009, 10: Q02014. doi:10.1029/2008GC002149. 引文使用过时参数coauthors (帮助)
^ (英文)Is the Gulf's Origin Heaven Sent? by Michael S. Stanton Explorer magazine article "Is the Gulf's Origin Heaven Sent?"
^ (英文)Jones AP, Price GD, Price NJ, DeCarli PS, Clegg RA. Impact induced melting and the development of large igneous provinces. Earth and Planetary Science Letters. 2002, 202 (3): 551–561. doi:10.1016/S0012-821X(02)00824-5.
^ 85.085.185.285.385.4 (英文)White RV. Earth’s biggest 'whodunnit': unravelling the clues in the case of the end-Permian mass extinction (PDF). Phil. Trans. Royal Society of London. 2002, 360: 2963–2985 [2008-01-12]. doi:10.1098/rsta.2002.1097.
^ (英文)AHager, Bradford H. Giant Impact Craters Lead To Flood Basalts: A Viable Model. CCNet 33/2001: Abstract 50470. 2001.
^ (英文)Hagstrum, Jonathan T. Large Oceanic Impacts As The Cause Of Antipodal Hotspots And Global Mass Extinctions. CCNet 33/2001: Abstract 50288. 2001.
^ (英文)Zhou, M-F., Malpas, J, Song, X-Y, Robinson, PT, Sun, M, Kennedy, AK, Lesher, CM & Keays, RR. A temporal link between the Emeishan large igneous province (SW China) and the end-Guadalupian mass extinction. Earth and Planetary Science Letters. 2002, 196 (3–4): 113–122. doi:10.1016/S0012-821X(01)00608-2.
^ (英文)Wignall, Paul B.; et al.. Volcanism, Mass Extinction, and Carbon Isotope Fluctuations in the Middle Permian of China. Science. 2009, 324 (5931): 1179–1182. doi:10.1126/science.1171956. 引文使用过时参数coauthors (帮助)
^ (英文)Mundil, R., Ludwig, K.R., Metcalfe, I. & Renne, P.R. Age and Timing of the Permian Mass Extinctions: U/Pb Dating of Closed-System Zircons. Science. 2004, 305 (5691): 1760–1763. doi:10.1126/science.1101012.
^ (英文)"Permian–Triassic Extinction - Volcanism"
^ (英文)Palfy J, Demeny A, Haas J, Htenyi M, Orchard MJ, & Veto I. Carbon isotope anomaly at the Triassic– Jurassic boundary from a marine section in Hungary. Geology. 2001, 29 (11): 1047–1050. doi:10.1130/0091-7613(2001)029%3C1047:CIAAOG%3E2.0.CO;2.
^ 93.093.193.2 (英文)Berner, R.A. Examination of hypotheses for the Permo-Triassic boundary extinction by carbon cycle modeling. Proceedings of the National Academy of Sciences. 2002, 99: 32095199. PMID 11917102. doi:10.1073/pnas.032095199.
^ 94.094.1 (英文)Dickens GR, O'Neil JR, Rea DK & Owen RM. Dissociation of oceanic methane hydrate as a cause of the carbon isotope excursion at the end of the Paleocene. Paleoceanography. 1995, 10 (6): 965–71. doi:10.1029/95PA02087.
^ (英文)Schrag, D.P., Berner, R.A., Hoffman, P.F., and Halverson, G.P. On the initiation of a snowball Earth. Geochemistry Geophysics Geosystems. 2002, 3 (6 doi=10.1029/2001GC000219): 1036. (原始内容存档于2008-04-21). Preliminary abstract at Schrag, D.P. On the initiation of a snowball Earth. Geological Society of America. June 2001.
^ 96.096.1 (英文)Benton, M.J.; Twitchett, R.J. How to kill (almost) all life: the end-Permian extinction event. Trends in Ecology & Evolution. 2003, 18 (7): 358–365. doi:10.1016/S0169-5347(03)00093-4. 引文使用过时参数coauthors (帮助)
^ (英文)Dickens GR. The potential volume of oceanic methane hydrates with variable external conditions. Organic Geochemistry. 2001, 32 (10): 1179–1193. doi:10.1016/S0146-6380(01)00086-9.
^ (英文)Reichow MK, Saunders AD, White RV, Pringle MS, Al'Muhkhamedov AI, Medvedev AI & Kirda NP. 40Ar/39Ar Dates from the West Siberian Basin: Siberian Flood Basalt Province Doubled. Science. 2002, 296 (5574): 1846–1849. doi:10.1126/science.1071671.
^ (英文)Holser WT, Schoenlaub H-P, Attrep Jr M, Boeckelmann K, Klein P, Magaritz M, Orth CJ, Fenninger A, Jenny C, Kralik M, Mauritsch H, Pak E, Schramm J-F, Stattegger K & Schmoeller R. A unique geochemical record at the Permian/Triassic boundary. Nature. 1989, 337 (6202): 39–44. doi:10.1038/337039a0.
^ (英文)Dobruskina IA. Phytogeography of Eurasia during the early Triassic. Palaeogeography, Palaeoclimatology, Palaeoecology. 1987, 58 (1-2): 75–86. doi:10.1016/0031-0182(87)90007-1.
^ (英文)Newell ND. An Outline History of Tropical Organic Reefs (PDF). American Museum novitates. 1971, 2465: 1–37 [2007-11-03].
^ (英文)McRoberts, C.A., Furrer, H., Jones, D.S. Palaeoenvironmental interpretation of a Triassic– Jurassic boundary section from western Austria based on palaeoecological and geochemical data.. Palaeogeography Palaeoclimatology Palaeoecology. 1997, 136 (1-4): 79– 95. doi:10.1016/S0031-0182(97)00074-6.
^ (英文)Wignall PB & Twitchett RJ. Permian–Triassic sedimentology of Jameson Land, East Greenland: Incised submarine channels in an anoxic basin. Journal of the Geological Society. 2002, 159 (6): 691–703. doi:10.1144/0016-764900-120.
^ 104.0104.1 (英文)Monastersky, R., Oxygen starvation decimated Permian oceans, Science News, May 25, 1996
^ 105.0105.1 Zhang R, Follows, MJ, Grotzinger, JP, & Marshall J. Could the Late Permian deep ocean have been anoxic?. Paleoceanography. 2001, 16 (3): 317–329. doi:10.1029/2000PA000522. (原始内容存档于2008-04-21).
^ (英文)Kump LR, Pavlov A, & Arthur MA. Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia. Geology. 2005, 33 (5): 397–400. doi:10.1130/G21295.1.
^ (英文)Grice K, Cao C, Love GD, Bottcher ME, Twitchett RJ, Grosjean E, Summons RE, Turgeon SC, Dunning W & Yugan J. Photic Zone Euxinia During the Permian–Triassic Superanoxic Event. Science. 2005, 307 (5710): 706–709. doi:10.1126/science.110432.
^ (英文)The Permian - Palaeos 互联网档案馆的存檔,存档日期2007-04-28.
延伸阅读
(英文).mw-parser-output .smallcaps{font-variant:small-caps}.mw-parser-output .nocaps{text-transform:lowercase}
Over, Jess (editor), Understanding Late Devonian and Permian–Triassic Biotic and Climatic Events, (Volume 20 in series Developments in Palaeontology and Stratigraphy (2006). The state of the inquiry into the extinction events.
(英文)
Sweet, Walter C. (editor), Permo–Triassic Events in the Eastern Tethys : Stratigraphy Classification and Relations with the Western Tethys (in series World and Regional Geology)
外部連結
- 專門性:
- 二疊紀-三疊紀滅絕事件:化石證據、滅亡物種、滅絕原因與分析
- 二疊紀-三疊紀滅絕事件:海洋、陸地動物與植物的滅絕,碳、氧、鍶同位素的變化,年代測定,時間與過程模式
- 與二疊紀末滅絕事件有關的南極洲冰層證據
- 生物史上的五次滅絕事件
- 西伯利亞暗色岩的年代、規模、成因、與影響
- 新聞性:
- 科學網:2.5億年前兩大物種滅絕之謎獲突破性進展
- BBC:二疊紀末滅絕事件的複雜原因
- BBC2:在2.5億年前,地球上的生物幾乎滅亡
- SpaceRef:南極洲冰層下的隕石坑(影像由俄亥俄州立大學提供)
- Science Daily:全球暖化使大氣層充滿硫化氫,並導致二疊紀末滅絕事件
- Science Daily:南極洲的冰層下發現疑似隕石坑的大坑洞
SPACE.com:南極洲發現地表最大的隕石坑,可能是地質年代中最嚴重的生物滅絕有關]- Space.com:地層中的二疊紀末生物滅絕證據
- NAI:二疊紀滅絕事件是否由撞擊事件導致?
生物集群灭绝事件 |
|---|
規模稍小的滅絕事件 ↓埃迪卡拉纪末期? ↓劳阶 ↓托阿尔阶 ↓阿普第阶 ↓森諾曼期-土侖期 ↓中新世 ↓石炭纪雨林崩溃 ↓波托米阶末期 ↓寒武纪-奥陶纪 ↓奧陶紀-志留紀 ↓泥盆紀後期 ↓二疊紀-三疊紀 ↓三疊紀-侏羅紀 ↓白堊紀-第三紀 ↓全新世、第四紀 大型滅絕事件 埃迪卡拉纪 侏羅紀 古近纪 新元古代 古生代 中生代 新生代 │ −600 │ −550 │ −500 │ −450 │ −400 │ −350 │ −300 │ −250 │ −200 │ −150 │ −100 │ −50 │ 0 距今百萬年前 |



Comments
Post a Comment