 Mixing
Mixing
蜥腳下目
| 蜥腳下目 化石时期:晚三疊紀到晚白堊紀,210–66 Ma PreЄ Є O S D C P T J K Pg N | |
|---|---|
 | |
| 路氏迷惑龍骨架模型,位于卡内基自然史博物馆 | |
科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 爬行纲 Reptilia |
| 演化支: | 恐龙总目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 演化支: | †板龙类 Plateosauria |
| 演化支: | †大脚类 Massopoda |
| 演化支: | †蜥脚形类 Sauropodiformes |
| 演化支: | †近蜥龍類 Anchisauria |
| 下目: | †蜥腳下目 Sauropoda Marsh,1878 |
演化支[1] | |
| |
異名 | |
| |
蜥腳下目(学名:Sauropoda)又稱蜥腳亞目、龍腳下目、真蜥腳亞目,在希臘文裡意為「有蜥蜴般的腳」,是蜥臀目的一個下目,是由奧塞內爾·查利斯·馬什(Othniel Charles Marsh)在1878年建立[2]。
蜥腳類恐龍具有小型頭部、長頸部、長尾巴、以及粗壯的四肢。牠們是目前已知陸地上出現過的最巨大動物,包括許多知名的屬,如迷惑龍(原名為雷龍)、腕龍、梁龍等。蜥腳類恐龍首次出現於晚三疊紀,牠們當時的外表類似原蜥腳下目恐龍。到了侏儸紀晚期(1億5000萬年前),蜥腳類恐龍的分布廣泛,尤其是梁龍科與腕龍科。只有泰坦巨龍類存活到白堊紀晚期,但牠們幾乎分佈於全球。然而,泰坦巨龍類與其他非鳥類的恐龍在白堊紀-第三紀滅絕事件中滅絕了。蜥腳下目的化石在各大陸都有發現,包含南極洲。
但是,很少發現完整的蜥腳類化石。許多物種,尤其是最大型的物種,僅發現相關聯或是非天然狀態的骨頭。許多接近完整的標本都缺乏頭部、尾巴末端、以及四肢。蜥腳下目和原蜥腳亞目都屬於蜥腳形亞目,他們和絕大多數恐龍都屬於蜥臀目。
目录
1 體徵
1.1 體型
1.2 四肢與腳掌
1.3 氣囊
1.4 身體裝甲
2 古生物學
2.1 生態位
2.2 群體行動與親代養育
2.3 步態
2.4 頭部與頸部姿勢
2.5 足跡化石與移動方式
2.6 體型的演化
3 發現歷史
4 分類學
4.1 分類
5 參考資料
6 延伸閱讀
7 外部連結
體徵
蜥腳類恐龍是草食性四足動物,通常擁有長頸部與匙狀牙齒。牠們擁有小型頭部、巨大身體、通常擁有長尾巴。牠們的後肢粗壯、筆直,後腳掌有五個腳趾,只有內側三、四趾具有趾爪。與後肢相比,前肢較為修長,前腳掌只有拇指有大型指爪。大部分科普書籍的想像圖,在這些無趾爪的位置加上蹄狀趾爪。在蜥腳類恐龍之中,近側尾椎有高度可鑑定性[3]。
蜥腳類恐龍的部分背椎移到頸部的位置,導致頸部至少有12節脊椎,延長了頸部長度。脊椎的神經棘形狀可容納韌帶,以協助抬高頸部與支撐頭部重量。尾椎至少有44節。柱狀四肢演化成可支撐龐大身體重量,踝部有軟骨可緩衝、避震。前肢有五根指頭,但指頭骨頭數量減少。牠們以腳趾行走,腳掌離地且有肉墊支撐重量。大型鼻開孔位於頭顱骨的後段。而恥骨或許可能支撐大型腸道[4]。
體型
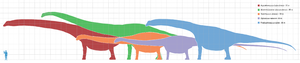
數種巨型蜥腳下目恐龍的體型比較圖。分別為:阿根廷龍(紅)、馬門溪龍(綠)、超龍(橘)、梁龍(紫)、富塔隆柯龍(藍)
蜥腳下目的最明顯特徵是牠們的體型。即使是侏儒型的蜥腳類恐龍,例如歐羅巴龍的身長可能達5到6公尺,也是牠們所處生態系統中最大的動物之一。唯一可以在體型上與牠們匹敵的是鬚鯨科動物,例如藍鯨。但與鯨魚不同的是,蜥腳類恐龍全都生存在陸地上。有些蜥腳類恐龍的頭部維持在低高度,例如梁龍科;而其他蜥腳類恐龍則維持在很高的高度,例如圓頂龍。
蜥腳類恐龍的身體外形並沒有其他恐龍多樣化,原因可能是體型限制,但牠們仍擁有許多種類。有些蜥腳類恐龍擁有極長的尾巴,例如梁龍科,牠們可能將尾巴當作鞭子般揮打,末端可產生音爆[5],或是造成掠食動物的傷害[6]。在化石較為完整的蜥腳類恐龍中,超龍可能是最長的恐龍,身長可達33到34公尺[7],但其他如地震龍、舊紀錄保持者梁龍也擁有極長的身長。易碎雙腔龍(Amphicoelias fragillimus)只發現一個脊椎骨,但身長可能達到55到60公尺[8],比藍鯨還長,但易碎雙腔龍的唯一化石卻在敘述不久後遺失了。而目前最長的陸地脊椎動物是網紋蟒,身長只有8.7公尺[9]。

柏林洪堡博物館的長頸巨龍(原名布氏腕龍)骨架模型,是目前全球最高的骨架模型
其他蜥腳類恐龍,例如腕龍科,擁有高肩膀與極長頸部,使牠們的身高非常高。波塞東龍可能是最高的恐龍,身高可達18公尺。而原本的頸部最長恐龍為馬門溪龍,身長有22到26公尺。而目前最高的動物是長頸鹿,身高只有4.8到5.5公尺。
其他體型特別大的蜥腳類恐龍包含:阿根廷龍的體重為80到100公噸之間,可能是最重的恐龍;富塔隆柯龍、潮汐龍、安第斯龍、南極龍、銀龍等大型恐龍的體型接近阿根廷龍。有些泰坦巨龍類可能更重,如巨體龍的體重可能為175到220公噸之間,但證據很少。而目前最大的陸地動物為非洲象,體重不超過10公噸。
最小型的蜥腳類恐龍包含:歐姆殿龍(4公尺)、侏儒種泰坦巨龍類馬扎爾龍(5.3公尺)、以及侏儒種腕龍科的歐羅巴龍(6.2公尺)[10]。發現於德國的歐羅巴龍,其小型體型可能是因為島嶼環境隔離而造成的。梁龍科的短頸潘龍擁有非常短的頸部,是梁龍科中最短的物種。其他蜥腳類恐龍的頸部可生長到背長的四倍,但短頸潘龍的頸部比牠們的身體還短。
四肢與腳掌
蜥腳類恐龍是群體型巨大的四足動物,因此四肢能夠承受相當大的重量。蜥腳類恐龍與現代大型四足動物(例如大象)的前腳結構不同。牠們的前腳掌骨頭以垂直方式排列、接觸地面;而大象的前腳掌骨頭往兩側稱開,形成寬廣的腳掌,手指骨頭縮短。原始蜥腳類恐龍(例如火山齒龍、巨腳龍)的手部骨頭仍往兩側稱開,具有腳指[11]。真蜥腳類的前腳掌只有蹄狀指爪,無法看到腳指。蜥腳類恐龍的後腳掌寬廣,大部分物種只有三個腳趾[12]。
蜥腳類恐龍的掌骨以半圓形、垂直地面方式排列,所以牠們的前腳足跡呈馬蹄狀。根據足跡化石,蜥腳類的前腳掌沒有肉墊,與大象不同[11]。大部分蜥腳類只有拇指具有明顯的指爪,功能仍未知。梁龍科的拇指指爪最大;而腕龍科的拇指指爪最小,某些足跡化石甚至無法辨認出拇指指爪[13]。
除了早期物種(如詹尼斯龍),泰坦巨龍類沒有拇指指爪。泰坦巨龍類是最特殊的蜥腳類恐龍,除了沒有指爪,前腳掌的指骨都已退化、消失。泰坦巨龍類的前腳掌只有柱狀的掌骨[14]。
根據葡萄牙發現的足跡化石,至少某些蜥腳類恐龍(可能屬於腕龍科)的前腳掌的底部、側邊,覆蓋者小型的長刺狀鱗片[15]。泰坦巨龍類的掌骨底部(接觸地面端)的形狀寬廣、呈方形,某些標本的前腳掌保存了軟組織,顯示泰坦巨龍類的前腳掌可能具有肉墊[14]。
Matthew Bonnan提出蜥腳類恐龍的四肢骨頭,在成長的過程中會保持大致的形狀,長寬高以相等的程度增加。業餘科學家Jim Schmidt認為四肢以柱狀方式直立的動物,四肢骨頭會採取這種生長方式。大部分脊椎動物的四肢骨頭在成長過程中,長寬高的成長幅度不一致,造成形狀上的改變,以與增加的體重相符[16][17]。
氣囊
如同獸腳亞目與鳥類,蜥腳類恐龍的脊椎內部具有複雜的凹處、洞孔、空室,生前可能包含氣囊,類似鳥類的呼吸系統。早期蜥腳類恐龍只有頸椎具有氣腔,新蜥腳類的薦椎也出現氣腔。梁龍超科與泰坦巨龍類也個別發展出前段尾椎的氣腔[18]。
在蜥腳下目的研究歷史早期,古生物學家就已辨認出這些動物具有氣腔。在19世紀晚期命名的鳥面龍(Ornithopsis),曾經因為脊椎的空腔,而被認為是種翼龍類[19]。

葡萄園龍,一種身體覆蓋裝甲的泰坦巨龍類
身體裝甲
有些蜥腳類恐龍擁有由皮內成骨形成的裝甲(例:薩爾塔龍、葡萄園龍)。有些屬的背上擁有尖刺(如奧古斯丁龍),而其中幾屬擁有尾槌(蜀龍)。
古生物學
生態位
最初發現蜥腳類恐龍的化石時,許多科學家將牠們與現代鯨魚互相比較、參照。19世紀到20世紀早期的大部分相關研究,認為這些史前大型爬行動物的體型過大,無法在陸地上行動,因此認為牠們應該是水生動物。在1970年代以前,大部分古生物想像圖,將蜥腳類恐龍描繪成完全水生或半水生動物[20]。在1950年代,開始有古生物學家質疑蜥腳類恐龍是否是水生動物,如果牠們的身體沉浸於水面下,極大壓力將造成肺臟與心臟的傷害[21]。此外,大部分早期研究只注意到蜥腳類恐龍的巨大體型,而忽略牠們骨骼內部的氣囊空間。在1878年,古生物學家愛德華·德林克·科普曾提到蜥腳類恐龍的骨骼結構類似浮筒。
在1970年代,開始有古生物學家研究蜥腳類恐龍的多氣囊骨骼、水生生活方式之間的關係。羅伯特·巴克等人根據蜥腳類恐龍的多氣囊骨骼、沈積學與生物力學的證據,而提出蜥腳類恐龍其實是主要生存於陸地的動物。在2004年,D.M. Henderson提出蜥腳類恐龍的骨骼具有非常多氣囊空間,如果牠們進入水體時,浮力會讓牠們浮在水面、無法完全沉浸於水面之下。另一方面,由於牠們的身體不會完全位於水面下方,巨大壓力不會壓迫牠們的內臟器官[20]。
過去曾發現數個蜥腳類恐龍的足跡化石,只有前肢的足跡,被認為是蜥腳類恐龍在水中游泳的證據。D.M. Henderson提出,這可能是長前肢的蜥腳類恐龍(例如大鼻龍類)在淺水前進時,用長前肢在水底移動,而較短後肢不接觸水底[20]。但由於蜥腳類恐龍的身體外形,牠們在水面浮動時會很搖晃、不穩定。D.M. Henderson認為,當蜥腳類恐龍在水中以長前肢移動時,牠們將難以控制方向、身體不穩定,類似喝醉的船夫[20]。
目前蜥腳類恐龍不再被認為是水生或半水生動物,近年證據顯示牠們生存於潮濕或水岸環境。蜥腳類的足跡化石常被發現於水岸環境或泛濫平原的沉積層,而蜥腳類的骨骼化石常被發現於潮濕沉積層、或是帶有水生生物的化石[20]。
群體行動與親代養育

群體行動的阿拉莫龍想像圖
根據許多屍骨層與足跡化石,顯示蜥腳類恐龍是群居動物。但是不同種類的蜥腳類恐龍,構成群體的規模不一。以一個發現於阿根廷侏羅紀中期地層的屍骨層為例,當中的化石由成年個體、幼年個體所構成,來自於不同年齡層。而根據其他屍骨層、足跡化石,許多蜥腳類恐龍的群體依年齡層而構成,成年個體與幼年個體分別獨自行動。目前已知會以不同年齡層而個別群體行動的的物種,包含:阿拉莫龍、巧龍、以及某些梁龍科[22]。
在2009年,一群科學家試圖研究蜥腳類恐龍為何會以不同年齡層來群體行動。根據牙齒的磨損程度顯示,成年、幼年蜥腳類恐龍會以不同植物為食。研究人員推測,成年、幼年蜥腳類恐龍分別以群體行動,是尋求不同種類的食物來源。而成年與幼年個體的巨大差距,也可能影響牠們攝取植物的方式,並使牠們採取分別群體行動的方式[22]。

一個幼年梁龍科骨架模型
由於剛孵化的幼體,可能會與成年個體共同相處一段時間,而且蜥腳類恐龍可能是早熟性動物。研究人員推測,那些會採取不同年齡層分開行動的物種,不會有親代養育的行為[22]。而會採取成年、幼年個體群體行動的物種,可能會有親代養育的行為,以照顧剛孵化的幼體直到成年[23]。
目前古動物學家還不清楚哪些物種會採成年、幼年個體集體行動,哪些物種會採取不同年齡層分開行動。需要更多的群體行動證據,才能清楚研究出蜥腳類恐龍的群居行為模式[22]。
步態

以後腳站立的梁龍想像圖。由查爾斯·耐特在1911年繪製
在非常早期的時候,亨利·費爾費爾德·奧斯本(Henry Fairfield Osborn)與其他人便已假設蜥腳類恐龍能以後肢站立,使用牠們的尾巴充當三腳架的第三隻腳(有點類似袋鼠)[24]。美國自然歷史博物館的一個重龍著名骨架模型,便是採用這個假設而重建。一個2005年的研究假設,如果蜥腳類恐龍已適應偶爾以二足方式站立,那牠們的前肢腳掌應該會有因為壓力性骨折(Stress fracture)。然而,在檢驗過眾多的蜥腳類骨骸後,沒有發現相關的證據[25]。

以三腳架方式站立的重龍骨架模型。位於紐約美國自然歷史博物館
如果蜥腳類恐龍以三角架方式站立,應該會有巨大的重量施加在尾巴的脈棘(Haemal spine)上。隨者蜥腳類恐龍的成長,牠們的體重也隨之增加,當牠們以後腳站立時,這些脈棘會承受越來越多的重量,直到某些脈棘發生壓力性骨折,這將讓蜥腳類恐龍以後腳站立時感到痛苦,而因此必須改採四足方式。當蜥腳類恐龍過重而無法後腳站立時,牠們可能發展出較為安全的方施以避免用後腳站立。在蜥腳類恐龍的尾椎上曾發現骨折的脈棘。
在2009年,Heinrich Mallison研究不同蜥腳類恐龍以三腳架方式站立的可能性。他發現某些被認為是以後肢站立的特徵,例如泰坦巨龍類的左右腳間距寬,其實並不代表牠們能夠用後肢站立。Mallison分別指出,泰坦巨龍類的脊柱關節靈活,如果以三腳架方式站立,脊柱的靈活關節會使牠們無法穩定站立,除非背部有大量肌肉輔助。而腕龍科的身體重心位在身體前半段,也使牠們無法以三腳架方式站立[26]。梁龍科似乎較有可能以三腳架方式站立。梁龍科的頸部關節靈活、身體重心位在臀部、骨盆結構穩固、以及尾椎的形狀,可能允許尾巴負擔部分的重量。Mallison推測,梁龍科比大象更適合偶爾以後腳站立。Mallison更主張,當牠們偶爾以後肢站立時,四肢骨頭並不會發生壓力性骨折[26]。
頭部與頸部姿勢
長期以來,蜥腳類是否能夠高舉頭部、頸部,或是頸部維持在水平的姿勢,是最常提起的爭論之一。某些科學家們提出質疑,如果蜥腳類恐龍能夠高舉頸部,以高層的樹葉為食,牠們的心臟是否能夠維持足夠的血壓,供應血液至腦部。一項2009年的研究顯示,如果蜥腳類的頸部高舉,將血液輸送到頭部,會消耗大量的能量,佔去牠們日常攝取能量的一半以上[27]。此外,心臟若將血液輸送至高舉的頭部,血壓將高達700毫米汞柱,牠們的心臟大小必須是鯨魚的15倍[28]。有學者主張,梁龍、迷惑龍等蜥腳類恐龍進食時,身體的位置保持不動,藉由長頸部的左右移動,以大範圍的低矮植被為食[29]。
抱持反對意見的學者,認為蜥腳類能夠以S狀方式高舉頸部。在2009年,一個研究提出所有四足類都能夠將頭頸部高舉,以警戒四周的環境。研究人員也發現,梁龍的頸部可以下垂至45度[30][31]。

一個蜥腳類恐龍的足跡化石
足跡化石與移動方式
目前已在許多大陸發現蜥腳類的足跡化石。古動物學家藉由這些足跡化石,可以研究蜥腳類的前肢、後肢功能。蜥腳類的前肢足跡小於後肢足跡,前肢足跡的形狀常為新月狀[32]。
根據左右足跡的間距,蜥腳類恐龍的足跡化石可分為三種型態,分別為:狹窄間距、中等間距、寬間距,可推論出不同的移動方式[32]。一個2004年的研究,發現進階型蜥腳類恐龍的足跡間距可歸納出不同型態,與演化上的分類相符合。大部分蜥腳類恐龍的足跡間距狹窄,前肢拇指的指爪痕跡明顯。腕龍科、原始巨龍形類的足跡間距、拇指指爪痕跡中等,顯示出牠們的演化趨勢。原始泰坦巨龍類的左右足跡間距更寬,仍保有拇指指爪。進階型泰坦巨龍類的左右足跡間距最寬,前肢的拇指指爪已消失[33]。
科學家們過去一度認為蜥腳龍類必須棲息在湖泊或沼澤中藉水漂浮。不過在找到化石足跡後,就推翻了這個說法。蜥腳類的足跡寬度間隔狹窄,證明牠們的四肢位於身體正下方,而非像蜥蜴與烏龜般向兩側張開[4]。
體型的演化
為何蜥腳類恐龍會演化出巨大的體型,目前已有數個相關理論。在蜥腳類恐龍的早期演化階段,就有演化出巨大體型的趨勢,最早可追溯至三疊紀晚期。肯尼思·卡彭特(Kenneth Carpenter)推測,蜥腳類恐龍的祖先有體型巨大化的演化壓力[8]。
根據體型巨大的現代草食性哺乳動物(例如象及犀牛),顯示巨大的草食性動物會有較高的消化效率。由於大型的動物有較長的消化系統,食物會留在身體較長時間,容許牠們在低營養價值的食物來源下生存。牠們的小腸會有大量的發酵室,容許微生物積聚及發酵植物,協助消化。在蜥腳類的演化歷史中,化石主要發現於半乾燥及季節性乾焊的環境,而在旱季有著季節性的食物營養價值下降。以大部分晚侏羅紀的蜥腳類恐龍為例,牠們的生存環境是疏林草原,而現代巨大草食性動物生存在類似的乾燥環境,支持上述的說法。其他巨大體型的優勢則被認為是次要的,例如對掠食動物的相對抵抗力、低能量消耗、或較長的壽命等[8]。
發現歷史

圓頂龍想像圖,由John A. Ryder於1877年繪製,是第一個蜥腳類的重建圖

目前的圓頂龍骨架模型
已知最早的蜥腳類化石,是在英格蘭發現。這些零散的化石,最初被鑑定為其他動物。當時並不清楚這些動物的外表,以及與其他恐龍的演化關係。
第一個被科學方式研究的蜥腳類化石,是一顆牙齒。在1699年,Edward Lhuyd在英格蘭牛津市近郊發現一顆牙齒化石,命名為Rutellum implicatum[34]。在2002年,科學家發現這顆牙齒屬於蜥腳下目恐龍。但是,這個學名的建立早於林奈氏分類系統的建立,而且已經超過50年不曾被任何科學期刊使用,而且是個根據牙齒而被建立的學名,因此本身也是個無效的遺失名[35]。
在1841年,理查·歐文(Richard Owen)根據稍早發現的大型零散骨頭,認為是種未知的大型海生爬行動物,是現代鱷魚的近親,於是命名為鯨龍(Cetiosaurus),意為「鯨魚蜥蜴」。同時,他將兩顆心臟形狀的牙齒,判斷是種未知的大型爬行動物,並命名為央齒龍(Cardiodon)。鯨龍與央齒龍成為最早被科學方式敘述、命名的蜥腳類恐龍。但在1842年,理查·歐文建立恐龍(Dinosauria)一詞,並當時沒有包含鯨龍與央齒龍[36]。
在1850年,吉迪恩·曼特爾(Gideon Mantell)重新研究數個被歸類於鯨龍的骨頭,他發現其腿部骨頭具有髓腔,這是種陸地動物的特徵。曼特爾將這些骨頭建立為畸形龍(Pelorosaurus),屬於恐龍。但是曼特爾並沒有發現畸形龍與鯨龍的相似處[19]。
在1870年,哈利·絲萊(Harry Seeley)鑑定一些脊椎骨,他發現這些脊椎屬於一種大型動物,內部具有氣囊。由於當時只有鳥類與翼龍目的骨頭具有氣囊,哈利·絲萊將這些脊椎命名為鳥面龍(Ornithopsis),意為「鳥的面孔」,並歸類於翼龍目[19]。
隨者更完整的鯨龍化石的出土,約翰·菲力普斯(John Phillips)在1871年提出鯨龍是畸形龍的近親,也屬於恐龍[37]。自從1870年代開始,美國發現更多、更完整的蜥腳類化石(尤其是迷惑龍、圓頂龍),古生物學界才拼湊出這些大型爬行動物的外形。在1877年,John A. Ryder繪製圓頂龍的重建圖,這是第一個蜥腳類恐龍的重建圖[38]。同樣在1877年,理查德·萊德克(Richard Lydekker)根據在印度發現的零散脊椎,命名為泰坦巨龍(Titanosaurus),這是首次在南方大陸發現的蜥腳類恐龍[19]。
在1878年,奧塞內爾·查利斯·馬什(Othniel Charles Marsh)發現了當時最完整的蜥腳類化石,並命名為梁龍。馬什同時建立蜥腳亞目(Sauropoda),意為「蜥蜴的腳」,包含鯨龍、梁龍、以及牠們的近親,以有別於其他已知的恐龍分類單元[19]。
分類學

數種大鼻龍類恐龍,由左到右依序為:
圓頂龍、腕龍、長頸巨龍、盤足龍
在最近幾年,蜥腳下目的分類大致上是固定的,但還是有一些科或屬的位置未確定,例如:盤足龍、簡棘龍、約巴龍、以及納摩蓋吐龍科。在某些版本分類法裡,並不包含火山齒龍科、鯨龍科、峨嵋龍科,因為牠們被認為是並系群,或複系群;某些版本也認為圓頂龍科是複系群,而不包含圓頂龍科。
分類
以下簡略演化樹是根據威爾森在2002年的研究[39]:
| 蜥腳下目 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
以下演化樹來自於迪亞戈·玻爾(Diego Pol)等人的2011年研究[40]:
| 蜥腳下目 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
參考資料
^ Holtz, Thomas R. Jr. Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages (PDF). Random House. 2012.
^ Marsh, O.C. (1878). "Principal characters of American Jurassic dinosaurs. Part I". American Journal of Science and Arts 16: 411-416
^ Tidwell, V., Carpenter, K. & Meyer, S. 2001. New Titanosauriform (Sauropoda) from the Poison Strip Member of the Cedar Mountain Formation (Lower Cretaceous), Utah. In: Mesozoic Vertebrate Life. D. H. Tanke & K. Carpenter (eds.). Indiana University Press, Eds. D.H. Tanke & K. Carpenter. Indiana University Press. 139-165.
^ 4.04.1 David Lambert, Darren Naish, Elizabeth Wyse. Encyclopedia of Dinosaurs and Prehistoric Life. Dorling Kindersley Publishers Ltd. 2002.
^ Peterson, Ivars. Whips and Dinosaur Tails. Science News. March 2000 [2007-07-07].
^ Bakker, Robert (1994). "The Bite of the Bronto" Earth 3:(6):26–33.
^ Lovelace, David M.; Hartman, Scott A.; and Wahl, William R. Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny. Arquivos do Museu Nacional. 2007, 65 (4): 527–544. 引文使用过时参数coauthors (帮助)
^ 8.08.18.2 Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131-138.存档副本 (PDF). [2008-01-08]. (原始内容 (PDF)存档于2007-12-02).
^ Murphy JC, Henderson RW. 1997. Tales of Giant Snakes: A Historical Natural History of Anacondas and Pythons. Krieger Pub. Co. 221 pp. ISBN 978-0-89464-995-0.
^ Martin Sander, P.; Mateus, Octávio; Laven, Thomas; Knötschke, Nils. Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur. Nature. 2006, 441 (7094): 739–41. PMID 16760975. doi:10.1038/nature04633.
^ 11.011.1 Bonnan, M.F. (2003). "The evolution of manus shape in sauropod dinosaurs: implications for functional morphology, forelimb orientation, and phylogeny." Journal of Vertebrate Paleontology, 23: 595-613.
^ Paul, G.S. (1987). "The science and art of restoring the life appearance of dinosaurs and their relatives - a rigorous how-to guide." Pp. 4-49 in Czerkas, S.J. and Olson, E.C. (eds.), Dinosaurs Past and Present Vol. II. Seattle: University of Washington Press.
^ Upchurch, P. Manus claw function in sauropod dinosaurs. Gaia. 1994, 10: 161–171.
^ 14.014.1 Apesteguía, S. (2005). "Evolution of the titanosaur metacarpus." Pp. 321-345 in Tidwell, V. and Carpenter, K. (eds.) Thunder-Lizards: The Sauropodomorph Dinosaurs. Indianapolis: Indiana University Press.
^ Milàn, J.; Christiansen, P.; Mateus, O. A three-dimensionally preserved sauropod manus impression from the Upper Jurassic of Portugal: implications for sauropod manus shape and locomotor mechanics. Kaupia. 2005, 14: 47–52.
^ Bonnan, M. F. Morphometric Analysis of Humerus and Femur Shape in Morrison Sauropods: Implications for Functional Morphology and Paleobiology. Paleobiology. 2004, 30 (3): 444–470. JSTOR 4096900.
^ Bonnan, Matthew F. Linear and Geometric Morphometric Analysis of Long Bone Scaling Patterns in Jurassic Neosauropod Dinosaurs: Their Functional and Paleobiological Implications. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. 2007, 290 (9): 1089. doi:10.1002/ar.20578.
^ Wedel, M.J. (2009). "Evidence for bird-like air sacs in Saurischian dinosaurs." Journal of Experimental Zoology, 311A: 18pp.
^ 19.019.119.219.319.4 Taylor, M.P. (In press). "Sauropod dinosaur research: a historical review." In Richard Moody, Eric Buffetaut, David M. Martill and Darren Naish (eds.), Dinosaurs (and other extinct saurians): a historical perspective. HTML abstract.
^ 20.020.120.220.320.4 Henderson, D.M. (2004). "Tipsy punters: sauropod dinosaur pneumaticity, buoyancy and aquatic habits." Proceedings of the Royal Society of London B, 71: S180–S183. doi:10.1098/rsbl.2003.0136
^ Kermack, K.A. A note on the habits of sauropods. Ann. Mag. Nat. Hist. 1951, 4: 830–832.
^ 22.022.122.222.3 Myers, T.S.; Fiorillo, A.R. Evidence for gregarious behavior and age segregation in sauropod dinosaurs. Palaeogeography, Palaeoclimatology, Palaeoecology. 2009, 274: 96–104.
^ Coria, R.A. On a monospecific assemblage of sauropod dinosaurs from Patagonia: implications for gregarious behavior. GAIA. 1994, 10: 209–213.
^ Osborn, H. F. A Skeleton of Diplodocus, Recently Mounted in the American Museum. Science. 1899, 10 (259): 870–4. PMID 17788971. doi:10.1126/science.10.259.870.
^ Rothschild BM, Molnar RE. Sauropod Stress Fractures as Clues to Activity. (编) Carpenter, Kenneth and Tidswell, Virginia (ed.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. 2005: 381–391. ISBN 978-0-253-34542-4.
^ 26.026.1 Mallison, H. (2009). "Rearing for food? Kinetic/dynamic modeling of bipedal/tripodal poses in sauropod dinosaurs." P. 63 in Godefroit, P. and Lambert, O. (eds), Tribute to Charles Darwin and Bernissart Iguanodons: New Perspectives on Vertebrate Evolution and Early Cretaceous Ecosystems. Brussels.
^ Raising the sauropod neck: it costs more to get less. Biol. Lett. June 2009, 5 (3): 317–9. PMC 2679936. PMID 19364714. doi:10.1098/rsbl.2009.0096.
^ Hearts, neck posture and metabolic intensity of sauropod dinosaurs. Proc. Biol. Sci. September 2000, 267 (1455): 1883–7. PMC 1690760. PMID 11052540. doi:10.1098/rspb.2000.1225.
^ Stevens, K.A.; Parrish, J.M. Neck posture and feeding habits of two Jurassic sauropod dinosaurs. Science. 1999, 284 (5415): 798–800. PMID 10221910. doi:10.1126/science.284.5415.798.
^ Taylor, M.P., Wedel, M.J., and Naish, D. (2009). "Head and neck posture in sauropod dinosaurs inferred from extant animals". Acta Palaeontologica Polonica 54 (2), 2009: 213-220abstract
^ Museums and TV have dinosaurs' posture all wrong, claim scientists. Guardian, 27 May 2009
^ 32.032.1 Riga, B.J.G. and Calvo, J.O. (2009). "A new wide-gauge sauropod track site from the Late Cretaceous of Mendoza, Neuquen Basin, Argentina." Paleontology, 52(3): 631-640.
^ Day, J.J.; Norman, D.B.; Gale, A.S.; Upchurch, P.; Powell, H.P. A Middle Jurassic dinosaur trackway site from Oxfordshire, UK. Palaeontology. 2004, 47: 319–348.
^ Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Gleditsch and Weidmann: London.
^ Delair, J.B.; Sarjeant, W.A.S. The earliest discoveries of dinosaurs: the records re-examined. Proceedings of the Geologists' Association. 2002, 113: 185–197. doi:10.1016/S0016-7878(02)80022-0.
^ Owen, R. (1842). "Report on British Fossil Reptiles." Part II. Report of the British Association for the Advancement of Science, Plymouth, England.
^ Phillips, J. (1871). Geology of Oxford and the Valley of the Thames. Oxford: Clarendon Press, 523 pp.
^ Osborn, H.F., and Mook, C.C. (1921). "Camarasaurus, Amphicoelias and other sauropods of Cope." Memoirs of the American Museum of Natural History, n.s. 3:247-387 and plates LX-LXXXV.
^ Wilson, J. A. (2002). "Sauropod dinosaur phylogeny: critique and cladistic analysis." Zoological Journal of the Linnean Society, 136: 217-276.
^ Cecilia Apaldetti, Ricardo N. Martinez, Oscar A. Alcober and Diego Pol. A New Basal Sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), Northwestern Argentina. PLoS ONE. 2011, 6 (11): e26964. doi:10.1371/journal.pone.0026964.
延伸閱讀
- Bob Strauss, 2008, Sauropods: The Biggest Dinosaurs that Ever Lived, The New York Times
- Kristina Curry Rogers and Jeffrey A. Wilson, 2005, The Sauropods: Evolution and Paleobiology, University of California Press, Berkeley, ISBN 978-0-520-24623-2
- Upchurch, P., Barrett, P.M. and Dodson, P. 2004. Sauropoda. In The Dinosauria, 2nd edition. D. Weishampel, P. Dodson, and H. Osmólska (eds.). University of California Press, Berkeley. pp. 259–322.
外部連結
(英文)蜥腳下目的詳述 - DinoData
(英文)蜥腳下目的特徵 - Palaeos
(英文)蜥腳下目的分類系統 - Thescelosaurus!
(英文)(德文)[https://web.archive.org/web/20091205215252/http://www.sauropod-dinosaurs.uni-bonn.de/deutsch/index_dt.htm 蜥腳下目的生理與演化
| |||||||||||||||||||||||||||||||||||||||||||




Comments
Post a Comment