 Mixing
Mixing
GABAA receptor
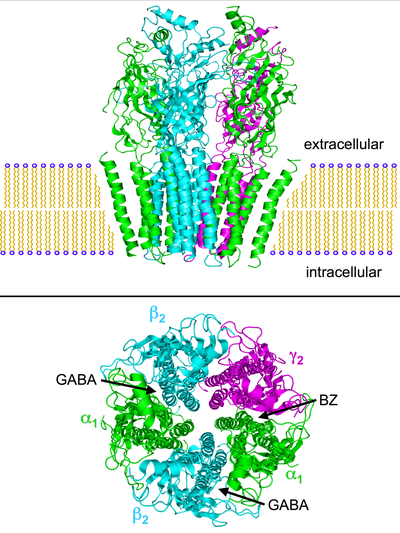
Structure of the nicotinic acetylcholine receptor (nAchR: PDB: 2BG9) which is very similar to the GABAA receptor.[1][2][3]Top: side view of the nAchR embedded in a cell membrane. Bottom: view of the receptor from the extracellular face of the membrane. The subunits are labeled according to the GABAA nomenclature and the approximate locations of the GABA and benzodiazepine (BZ) binding sites are noted (between the α- and β-subunits and between the α- and γ-subunits respectively).

Schematic structure of the GABAA receptor. Left: GABAA monomeric subunit embedded in a lipid bilayer (yellow lines connected to blue spheres). The four transmembrane α-helices (1–4) are depicted as cylinders. The disulfide bond in the N-terminal extracellular domain which is characteristic of the family of cys-loop receptors (which includes the GABAA receptor) is depicted as a yellow line. Right: Five subunits symmetrically arranged about the central chloride anion conduction pore. The extracellular loops are not depicted for the sake of clarity.
The GABAA receptor (GABAAR) is an ionotropic receptor and ligand-gated ion channel. Its endogenous ligand is γ-aminobutyric acid (GABA), the major inhibitory neurotransmitter in the central nervous system. Upon activation, the GABAA receptor selectively conducts Cl− through its pore. Cl- will flow out of the cell if the internal voltage is less than resting potential and Cl- will flow in if it is more than resting potential (i.e. -75 mV)[4]. This causes an inhibitory effect on neurotransmission by diminishing the chance of a successful action potential occurring. The reversal potential of the GABAA-mediated inhibitory postsynaptic potential (IPSP) in normal solution is −70 mV, contrasting the GABAB IPSP (-100 mV).
The active site of the GABAA receptor is the binding site for GABA and several drugs such as muscimol, gaboxadol, and bicuculline.[5] The protein also contains a number of different allosteric binding sites which modulate the activity of the receptor indirectly. These allosteric sites are the targets of various other drugs, including the benzodiazepines, nonbenzodiazepines, neuroactive steroids, barbiturates, alcohol (ethanol),[6]inhaled anaesthetics, and picrotoxin, among others.[7]
GABAA receptors occur in all organisms that have a nervous system. To a limited extent the receptors can be found in non-neuronal tissues. Due to their wide distribution within the nervous system of mammals they play a role in virtually all brain functions.
Contents
1 Target for benzodiazepines
2 Structure and function
2.1 Subunits
2.2 Distribution
3 Ligands
3.1 Types
3.2 Examples
3.3 Effects
3.4 Novel drugs
4 See also
5 References
6 Further reading
7 External links
Target for benzodiazepines
The ionotropic GABAA receptor protein complex is also the molecular target of the benzodiazepine class of tranquilizer drugs. Benzodiazepines do not bind to the same receptor site on the protein complex as the endogenous ligand GABA (whose binding site is located between α- and β-subunits), but bind to distinct benzodiazepine binding sites situated at the interface between the α- and γ-subunits of α- and γ-subunit containing GABAA receptors.[8][9] While the majority of GABAA receptors (those containing α1-, α2-, α3-, or α5-subunits) are benzodiazepine sensitive, there exists a minority of GABAA receptors (α4- or α6-subunit containing) which are insensitive to classical 1,4-benzodiazepines,[10] but instead are sensitive to other classes of GABAergic drugs such as neurosteroids and alcohol. In addition peripheral benzodiazepine receptors exist which are not associated with GABAA receptors. As a result, the IUPHAR has recommended that the terms "BZ receptor", "GABA/BZ receptor" and "omega receptor" no longer be used and that the term "benzodiazepine receptor" be replaced with "benzodiazepine site".[11]
In order for GABAA receptors to be sensitive to the action of benzodiazepines they need to contain an α and a γ subunit, between which the benzodiazepine binds. Once bound, the benzodiazepine locks the GABAA receptor into a conformation where the neurotransmitter GABA has much higher affinity for the GABAA receptor, increasing the frequency of opening of the associated chloride ion channel and hyperpolarising the membrane. This potentiates the inhibitory effect of the available GABA leading to sedative and anxiolytic effects.[citation needed]
Different benzodiazepines have different affinities for GABAA receptors made up of different collection of subunits, and this means that their pharmacological profile varies with subtype selectivity. For instance, benzodiazepine receptor ligands with high activity at the α1 and/or α5 tend to be more associated with sedation, ataxia and amnesia, whereas those with higher activity at GABAA receptors containing α2 and/or α3 subunits generally have greater anxiolytic activity.[12]Anticonvulsant effects can be produced by agonists acting at any of the GABAA subtypes, but current research in this area is focused mainly on producing α2-selective agonists as anticonvulsants which lack the side effects of older drugs such as sedation and amnesia.
The binding site for benzodiazepines is distinct from the binding site for barbiturates and GABA on the GABAA receptor, and also produces different effects on binding,[13] with the benzodiazepines increasing the frequency of the chloride channel opening, while barbiturates increase the duration of chloride channel opening when GABA is bound.[14] Since these are separate modulatory effects, they can both take place at the same time, and so the combination of benzodiazepines with barbiturates is strongly synergistic, and can be dangerous if dosage is not strictly controlled.
Also note that some GABAA agonists such as muscimol and gaboxadol do bind to the same site on the GABAA receptor complex as GABA itself, and consequently produce effects which are similar but not identical to those of positive allosteric modulators like benzodiazepines.[citation needed]
Structure and function

Schematic diagram of a GABAA receptor protein ((α1)2(β2)2(γ2)) which illustrates the five combined subunits that form the protein, the chloride (Cl−) ion channel pore, the two GABA active binding sites at the α1 and β2 interfaces, and the benzodiazepine (BDZ) allosteric binding site[1]
Structural understanding of the GABAA receptor was initially based on homology models, obtained using crystal structures of homologous proteins like Acetylcholine binding protein (AChBP) and nicotinic acetylcholine (nACh) receptors as templates.[15][16] The much sought structure of a GABAA receptor was finally resolved, with the disclosure of the crystal structure of human β3 homopentameric GABAA receptor.[17]
Whilst this was a major development, the majority of GABAA receptors are heteromeric and the structure did not provide any details of the benzodiazepine binding site. This was finally elucidated in 2018 by the publication of a high resolution cryo-EM structure of a human α1β2γ2 receptor bound with GABA and the neutral benzodiazepine flumazenil.[18]
GABAA receptors are pentameric transmembrane receptors which consist of five subunits arranged around a central pore. Each subunit comprises four transmembrane domains with both the N- and C-terminus located extracellularly. The receptor sits in the membrane of its neuron, usually localized at a synapse, postsynaptically. However, some isoforms may be found extrasynaptically.[19] The ligand GABA is the endogenous compound that causes this receptor to open; once bound to GABA, the protein receptor changes conformation within the membrane, opening the pore in order to allow chloride anions (Cl−) to pass down an electrochemical gradient. Because the reversal potential for chloride in most neurons is close to or more negative than the resting membrane potential, activation of GABAA receptors tends to stabilize or hyperpolarise the resting potential, and can make it more difficult for excitatory neurotransmitters to depolarize the neuron and generate an action potential. The net effect is typically inhibitory, reducing the activity of the neuron. However, depolarizing responses have been found to occur in response to GABA in immature neurons due to a modified Cl- gradient.[20] These depolarization events have shown to be key in neuronal development.[21] In the mature neuron, the GABAA channel opens quickly and thus contributes to the early part of the inhibitory post-synaptic potential (IPSP).[22][23]
The endogenous ligand that binds to the benzodiazepine site is inosine.[24][citation needed]
Subunits
GABAA receptors are members of the large pentameric ligand gated ion channel (previously referred to as "Cys-loop" receptors) super-family of evolutionarily related and structurally similar ligand-gated ion channels that also includes nicotinic acetylcholine receptors, glycine receptors, and the 5HT3 receptor. There are numerous subunit isoforms for the GABAA receptor, which determine the receptor's agonist affinity, chance of opening, conductance, and other properties.[25]
In humans, the units are as follows:[26]
- six types of α subunits (GABRA1, GABRA2, GABRA3, GABRA4, GABRA5, GABRA6)
- three βs (GABRB1, GABRB2, GABRB3)
- three γs (GABRG1, GABRG2, GABRG3)
- as well as a δ (GABRD), an ε (GABRE), a π (GABRP), and a θ (GABRQ)
There are three ρ units (GABRR1, GABRR2, GABRR3); however, these do not coassemble with the classical GABAA units listed above,[27] but rather homooligomerize to form GABAA-ρ receptors (formerly classified as GABAC receptors but now this nomenclature has been deprecated[28] ).
Distribution
GABAA receptors are responsible for most of the physiological activities of GABA in the central nervous system, and the receptor subtypes vary significantly. Subunit composition can vary widely between regions and subtypes may be associated with specific functions. The minimal requirement to produce a GABA-gated ion channel is the inclusion of an α and a β subunit.[29] The most common GABAA receptor is a pentamer comprising two α's, two β's, and a γ (α1β2γ2).[26] In neurons themselves, the type of GABAA receptor subunits and their densities can vary between cell bodies and dendrites.[30] GABAA receptors can also be found in other tissues, including leydig cells, placenta, immune cells, liver, bone growth plates and several other endocrine tissues. Subunit expression varies between 'normal' tissue and malignancies, as GABAA receptors can influence cell proliferation.[31]
| Isoform | Synaptic/Extrasynaptic | Anatomical location |
|---|---|---|
| α1β3γ2S | Both | Widespread |
| α2β3γ2S | Both | Widespread |
| α3β3γ2S | Both | Reticular thalamic nucleus |
| α4β3γ2S | Both | Thalamic relay cells |
| α5β3γ2S | Both | Hippocampal pyramidal cells |
| α6β3γ2S | Both | Cerebellar granule cells |
| α1β2γ2S | Both | Widespread, most abundant |
| α4β3δ | Extrasynaptic | Thalamic relay cells |
| α6β3δ | Extrasynaptic | Cerebellar granule cells |
| α1β2 | Extrasynaptic | Widespread |
| α1β3 | Extrasynaptic | Thalamus, hypothalamus |
| α1β2δ | Extrasynaptic | Hippocampus |
| α4β2δ | Extrasynaptic | Hippocampus |
| α3β3θ | Extrasynaptic | Hypothalamus |
| α3β3ε | Extrasynaptic | Hypothalamus |
Ligands

GABAA receptor and where various ligands bind.
A number of ligands have been found to bind to various sites on the GABAA receptor complex and modulate it besides GABA itself.[which?] A ligand can possess one or more properties of the following types. Unfortunately the literature often does not distinguish these types properly.
Types
Orthosteric agonists and antagonists: bind to the main receptor site (the site where GABA normally binds, also referred to as the "active" or "orthosteric" site). Agonists activate the receptor, resulting in increased Cl− conductance. Antagonists, though they have no effect on their own, compete with GABA for binding and thereby inhibit its action, resulting in decreased Cl− conductance.
First order allosteric modulators: bind to allosteric sites on the receptor complex and affect it either in a positive (PAM), negative (NAM) or neutral/silent (SAM) manner, causing increased or decreased efficiency of the main site and therefore an indirect increase or decrease in Cl− conductance. SAMs do not affect the conductance, but occupy the binding site.
Second order modulators: bind to an allosteric site on the receptor complex and modulate the effect of first order modulators.
Open channel blockers: prolong ligand-receptor occupancy, activation kinetics and Cl ion flux in a subunit configuration-dependent and sensitization-state dependent manner.[33]
Non-competitive channel blockers: bind to or near the central pore of the receptor complex and directly block Cl− conductance through the ion channel.
Examples
- Orthosteric agonists: GABA, gaboxadol, isoguvacine, muscimol, progabide, piperidine-4-sulfonic acid (partial agonist).
- Orthosteric antagonists: bicuculline, gabazine.
- Positive allosteric modulators: barbiturates, benzodiazepines, certain carbamates (ex. carisoprodol, meprobamate, lorbamate), thienodiazepines, alcohol (ethanol), etomidate, glutethimide, kavalactones,[34]meprobamate, quinazolinones (ex. methaqualone, etaqualone, diproqualone), neuroactive steroids,[35]niacin/niacinamide,[36]nonbenzodiazepines (ex. zolpidem, eszopiclone), propofol, stiripentol,[37]theanine,[citation needed]valerenic acid, volatile/inhaled anesthetics, lanthanum,[38] and riluzole.[39]
- Negative allosteric modulators: flumazenil, Ro15-4513, sarmazenil, amentoflavone, and zinc.[40]
- Second-order modulators: (−)‐epigallocatechin‐3‐gallate[citation needed].
- Non-competitive channel blockers: cicutoxin, oenanthotoxin, pentylenetetrazol, picrotoxin[citation needed], thujone, and lindane.
Effects
Ligands which contribute to receptor activation typically have anxiolytic, anticonvulsant, amnesic, sedative, hypnotic, euphoriant, and muscle relaxant properties. Some such as muscimol and the z-drugs may also be hallucinogenic.[citation needed] Ligands which decrease receptor activation usually have opposite effects, including anxiogenesis and convulsion.[citation needed] Some of the subtype-selective negative allosteric modulators such as α5IA are being investigated for their nootropic effects, as well as treatments for the unwanted side effects of other GABAergic drugs.[41]
Novel drugs
A useful property of the many benzodiazepine site allosteric modulators is that they may display selective binding to particular subsets of receptors comprising specific subunits. This allows one to determine which GABAA receptor subunit combinations are prevalent in particular brain areas and provides a clue as to which subunit combinations may be responsible for behavioral effects of drugs acting at GABAA receptors. These selective ligands may have pharmacological advantages in that they may allow dissociation of desired therapeutic effects from undesirable side effects.[42] Few subtype selective ligands have gone into clinical use as yet, with the exception of zolpidem which is reasonably selective for α1, but several more selective compounds are in development such as the α3-selective drug adipiplon. There are many examples of subtype-selective compounds which are widely used in scientific research, including:
CL-218,872 (highly α1-selective agonist)
bretazenil (subtype-selective partial agonist)
imidazenil and L-838,417 (both partial agonists at some subtypes, but weak antagonists at others)
QH-ii-066 (full agonist highly selective for α5 subtype)
α5IA (selective inverse agonist for α5 subtype)
SL-651,498 (full agonist at α2 and α3 subtypes, and as a partial agonist at α1 and α5
- 3-acyl-4-quinolones: selective for α1 over α3[43]
See also
- 4-Iodopropofol
- GABA receptor
- GABAB receptor
- GABAA-ρ receptor
- Gephyrin
- Glycine receptor
- GABAA receptor positive allosteric modulators
- GABAA receptor negative allosteric modulators
References
^ ab Richter L, de Graaf C, Sieghart W, Varagic Z, Mörzinger M, de Esch IJ, Ecker GF, Ernst M (March 2012). "Diazepam-bound GABAA receptor models identify new benzodiazepine binding-site ligands". Nature Chemical Biology. 8 (5): 455–64. doi:10.1038/nchembio.917. PMC 3368153. PMID 22446838..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output .citation q{quotes:"""""""'""'"}.mw-parser-output .citation .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .citation .cs1-lock-limited a,.mw-parser-output .citation .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .citation .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-ws-icon a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/4/4c/Wikisource-logo.svg/12px-Wikisource-logo.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-maint{display:none;color:#33aa33;margin-left:0.3em}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
^ Campagna-Slater V, Weaver DF (January 2007). "Molecular modelling of the GABAA ion channel protein". Journal of Molecular Graphics & Modelling. 25 (5): 721–30. doi:10.1016/j.jmgm.2006.06.001. PMID 16877018.
^ Sancar F, Ericksen SS, Kucken AM, Teissére JA, Czajkowski C (January 2007). "Structural determinants for high-affinity zolpidem binding to GABA-A receptors". Molecular Pharmacology. 71 (1): 38–46. doi:10.1124/mol.106.029595. PMC 2583146. PMID 17012619.
^ "Equilibrium Potentials". courses.washington.edu. Retrieved 2018-04-22.
^ Chua HC, Chebib M (2017). GABAA Receptors and the Diversity in their Structure and Pharmacology. Advances in Pharmacology (San Diego, Calif.). Advances in Pharmacology. 79. pp. 1–34. doi:10.1016/bs.apha.2017.03.003. ISBN 9780128104132. PMID 28528665.
^ Santhakumar V, Wallner M, Otis TS (May 2007). "Ethanol acts directly on extrasynaptic subtypes of GABAA receptors to increase tonic inhibition". Alcohol. 41 (3): 211–21. doi:10.1016/j.alcohol.2007.04.011. PMC 2040048. PMID 17591544.
^ Johnston GA (1996). "GABAA receptor pharmacology". Pharmacology & Therapeutics. 69 (3): 173–98. doi:10.1016/0163-7258(95)02043-8. PMID 8783370.
^ Sigel E (August 2002). "Mapping of the benzodiazepine recognition site on GABA(A) receptors". Current Topics in Medicinal Chemistry. 2 (8): 833–9. doi:10.2174/1568026023393444. PMID 12171574.
^ Akabas MH (2004). GABAA receptor structure-function studies: a reexamination in light of new acetylcholine receptor structures. International Review of Neurobiology. 62. pp. 1–43. doi:10.1016/S0074-7742(04)62001-0. ISBN 978-0-12-366862-2. PMID 15530567.
^ Derry JM, Dunn SM, Davies M (March 2004). "Identification of a residue in the gamma-aminobutyric acid type A receptor alpha subunit that differentially affects diazepam-sensitive and -insensitive benzodiazepine site binding". Journal of Neurochemistry. 88 (6): 1431–8. doi:10.1046/j.1471-4159.2003.02264.x. PMID 15009644.
^ Barnard EA, Skolnick P, Olsen RW, Mohler H, Sieghart W, Biggio G, Braestrup C, Bateson AN, Langer SZ (June 1998). "International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function". Pharmacological Reviews. 50 (2): 291–313. PMID 9647870.
^ Atack JR (August 2003). "Anxioselective compounds acting at the GABA(A) receptor benzodiazepine binding site". Current Drug Targets. CNS and Neurological Disorders. 2 (4): 213–32. doi:10.2174/1568007033482841. PMID 12871032.
^ Hanson SM, Czajkowski C (March 2008). "Structural mechanisms underlying benzodiazepine modulation of the GABA(A) receptor". The Journal of Neuroscience. 28 (13): 3490–9. doi:10.1523/JNEUROSCI.5727-07.2008. PMC 2410040. PMID 18367615.
^ Twyman RE, Rogers CJ, Macdonald RL (March 1989). "Differential regulation of gamma-aminobutyric acid receptor channels by diazepam and phenobarbital". Annals of Neurology. 25 (3): 213–20. doi:10.1002/ana.410250302. PMID 2471436.
^ Ernst M, Bruckner S, Boresch S, Sieghart W (November 2005). "Comparative models of GABAA receptor extracellular and transmembrane domains: important insights in pharmacology and function". Molecular Pharmacology. 68 (5): 1291–300. doi:10.1124/mol.105.015982. PMID 16103045.
^ Vijayan RS, Trivedi N, Roy SN, Bera I, Manoharan P, Payghan PV, Bhattacharyya D, Ghoshal N (November 2012). "Modeling the closed and open state conformations of the GABA(A) ion channel--plausible structural insights for channel gating". Journal of Chemical Information and Modeling. 52 (11): 2958–69. doi:10.1021/ci300189a. PMID 23116339.
^ Miller PS, Aricescu AR (August 2014). "Crystal structure of a human GABAA receptor". Nature. 512 (7514): 270–5. doi:10.1038/nature13293. PMC 4167603. PMID 24909990.
^ Zhu S, Noviello CM, Teng J, Walsh RM, Kim JJ, Hibbs RE (July 2018). "Structure of a human synaptic GABAA receptor". Nature. 559 (7712): 67–72. doi:10.1038/s41586-018-0255-3. PMC 6220708. PMID 29950725.
^ Wei W, Zhang N, Peng Z, Houser CR, Mody I (November 2003). "Perisynaptic localization of delta subunit-containing GABA(A) receptors and their activation by GABA spillover in the mouse dentate gyrus". The Journal of Neuroscience. 23 (33): 10650–61. PMID 14627650.
^ Ben-Ari Y, Cherubini E, Corradetti R, Gaiarsa JL (September 1989). "Giant synaptic potentials in immature rat CA3 hippocampal neurones". The Journal of Physiology. 416: 303–25. doi:10.1113/jphysiol.1989.sp017762. PMC 1189216. PMID 2575165.
^ Spitzer NC (March 2010). "How GABA generates depolarization". The Journal of Physiology. 588 (Pt 5): 757–8. doi:10.1113/jphysiol.2009.183574. PMC 2834934. PMID 20194137.
^ Olsen RW, DeLorey TM (1999). "Chapter 16: GABA and Glycine". In Siegel GJ, Agranoff BW, Fisher SK, Albers RW, Uhler MD. Basic neurochemistry: molecular, cellular, and medical aspects (Sixth ed.). Philadelphia: Lippincott-Raven. ISBN 978-0-397-51820-3.
^ Chen K, Li HZ, Ye N, Zhang J, Wang JJ (October 2005). "Role of GABAB receptors in GABA and baclofen-induced inhibition of adult rat cerebellar interpositus nucleus neurons in vitro". Brain Research Bulletin. 67 (4): 310–8. doi:10.1016/j.brainresbull.2005.07.004. PMID 16182939.
^ Yarom M, Tang XW, Wu E, Carlson RG, Vander Velde D, Lee X, Wu J (2016-08-01). "Identification of inosine as an endogenous modulator for the benzodiazepine binding site of the GABAA receptors". Journal of Biomedical Science. 5 (4): 274–80. doi:10.1007/bf02255859. PMID 9691220.
^ Cossart R, Bernard C, Ben-Ari Y (February 2005). "Multiple facets of GABAergic neurons and synapses: multiple fates of GABA signalling in epilepsies". Trends in Neurosciences. 28 (2): 108–15. doi:10.1016/j.tins.2004.11.011. PMID 15667934.
^ ab Martin IL and Dunn SMJ. GABA receptors A review of GABA and the receptors to which it binds. Tocris Cookson LTD.
^ Enz R, Cutting GR (May 1998). "Molecular composition of GABAC receptors". Vision Research. 38 (10): 1431–41. doi:10.1016/S0042-6989(97)00277-0. PMID 9667009.
^ Olsen RW, Sieghart W (January 2009). "GABA A receptors: subtypes provide diversity of function and pharmacology". Neuropharmacology. 56 (1): 141–8. doi:10.1016/j.neuropharm.2008.07.045. PMC 3525320. PMID 18760291.
^ Connolly CN, Krishek BJ, McDonald BJ, Smart TG, Moss SJ (January 1996). "Assembly and cell surface expression of heteromeric and homomeric gamma-aminobutyric acid type A receptors". The Journal of Biological Chemistry. 271 (1): 89–96. doi:10.1074/jbc.271.1.89. PMID 8550630.
^ Lorenzo LE, Russier M, Barbe A, Fritschy JM, Bras H (September 2007). "Differential organization of gamma-aminobutyric acid type A and glycine receptors in the somatic and dendritic compartments of rat abducens motoneurons". The Journal of Comparative Neurology. 504 (2): 112–26. doi:10.1002/cne.21442. PMID 17626281.
^ ten Hoeve AL (2012). GABA receptors and the immune system. Thesis, Utrecht University
^ Mortensen M, Patel B, Smart TG (January 2011). "GABA Potency at GABA(A) Receptors Found in Synaptic and Extrasynaptic Zones". Frontiers in Cellular Neuroscience. 6: 1. doi:10.3389/fncel.2012.00001. PMC 3262152. PMID 22319471.
^ Haseneder R, Rammes G, Zieglgänsberger W, Kochs E, Hapfelmeier G (September 2002). "GABA(A) receptor activation and open-channel block by volatile anaesthetics: a new principle of receptor modulation?". European Journal of Pharmacology. 451 (1): 43–50. doi:10.1016/S0014-2999(02)02194-5. PMID 12223227.
^ Hunter, A (2006). "Kava (Piper methysticum) back in circulation". Australian Centre for Complementary Medicine. 25 (7): 529.
^ (a) Herd MB, Belelli D, Lambert JJ (October 2007). "Neurosteroid modulation of synaptic and extrasynaptic GABA(A) receptors". Pharmacology & Therapeutics. 116 (1): 20–34. arXiv:1607.02870. doi:10.1016/j.pharmthera.2007.03.007. PMID 17531325.; (b) Hosie AM, Wilkins ME, da Silva HM, Smart TG (November 2006). "Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites". Nature. 444 (7118): 486–9. doi:10.1038/nature05324. PMID 17108970.; (c)Agís-Balboa RC, Pinna G, Zhubi A, Maloku E, Veldic M, Costa E, Guidotti A (September 2006). "Characterization of brain neurons that express enzymes mediating neurosteroid biosynthesis". Proceedings of the National Academy of Sciences of the United States of America. 103 (39): 14602–7. doi:10.1073/pnas.0606544103. PMC 1600006. PMID 16984997.; (d) Akk G, Shu HJ, Wang C, Steinbach JH, Zorumski CF, Covey DF, Mennerick S (December 2005). "Neurosteroid access to the GABAA receptor". The Journal of Neuroscience. 25 (50): 11605–13. doi:10.1523/JNEUROSCI.4173-05.2005. PMID 16354918.; (e) Belelli D, Lambert JJ (July 2005). "Neurosteroids: endogenous regulators of the GABA(A) receptor". Nature Reviews. Neuroscience. 6 (7): 565–75. doi:10.1038/nrn1703. PMID 15959466.; (f) Pinna G, Costa E, Guidotti A (June 2006). "Fluoxetine and norfluoxetine stereospecifically and selectively increase brain neurosteroid content at doses that are inactive on 5-HT reuptake". Psychopharmacology. 186 (3): 362–72. doi:10.1007/s00213-005-0213-2. PMID 16432684.; (g) Dubrovsky BO (February 2005). "Steroids, neuroactive steroids and neurosteroids in psychopathology". Progress in Neuro-Psychopharmacology & Biological Psychiatry. 29 (2): 169–92. doi:10.1016/j.pnpbp.2004.11.001. PMID 15694225.; (h) Mellon SH, Griffin LD (2002). "Neurosteroids: biochemistry and clinical significance". Trends in Endocrinology and Metabolism. 13 (1): 35–43. doi:10.1016/S1043-2760(01)00503-3. PMID 11750861.; (i) Puia G, Santi MR, Vicini S, Pritchett DB, Purdy RH, Paul SM, Seeburg PH, Costa E (May 1990). "Neurosteroids act on recombinant human GABAA receptors". Neuron. 4 (5): 759–65. doi:10.1016/0896-6273(90)90202-Q. PMID 2160838.; (j) Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM (May 1986). "Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor". Science. 232 (4753): 1004–7. doi:10.1126/science.2422758. PMID 2422758.; (k) Reddy DS, Rogawski MA (2012). "Neurosteroids — Endogenous Regulators of Seizure Susceptibility and Role in the Treatment of Epilepsy". In Noebels JL, Avoli M, Rogawski MA, et al. Jasper's Basic Mechanisms of the Epilepsies [Internet]. 4th edition. Bethesda (MD): National Center for Biotechnology Information (US).
^ Toraskar, Mrunmayee; Pratima R.P. Singh; Shashank Neve (2010). "STUDY OF GABAERGIC AGONISTS" (PDF). Deccan Journal of Pharmacology. 1 (2): 56–69.
^ Fisher JL (January 2009). "The anti-convulsant stiripentol acts directly on the GABA(A) receptor as a positive allosteric modulator". Neuropharmacology. 56 (1): 190–7. doi:10.1016/j.neuropharm.2008.06.004. PMC 2665930. PMID 18585399.
^ Boldyreva AA (October 2005). "Lanthanum potentiates GABA-activated currents in rat pyramidal neurons of CA1 hippocampal field". Bulletin of Experimental Biology and Medicine. 140 (4): 403–5. doi:10.1007/s10517-005-0503-z. PMID 16671565.
^ He Y, Benz A, Fu T, Wang M, Covey DF, Zorumski CF, Mennerick S (February 2002). "Neuroprotective agent riluzole potentiates postsynaptic GABA(A) receptor function". Neuropharmacology. 42 (2): 199–209. doi:10.1016/s0028-3908(01)00175-7. PMID 11804616.
^ Hosie AM, Dunne EL, Harvey RJ, Smart TG (April 2003). "Zinc-mediated inhibition of GABA(A) receptors: discrete binding sites underlie subtype specificity". Nature Neuroscience. 6 (4): 362–9. doi:10.1038/nn1030. PMID 12640458.
^ Dawson GR, Maubach KA, Collinson N, Cobain M, Everitt BJ, MacLeod AM, Choudhury HI, McDonald LM, Pillai G, Rycroft W, Smith AJ, Sternfeld F, Tattersall FD, Wafford KA, Reynolds DS, Seabrook GR, Atack JR (March 2006). "An inverse agonist selective for alpha5 subunit-containing GABAA receptors enhances cognition". The Journal of Pharmacology and Experimental Therapeutics. 316 (3): 1335–45. doi:10.1124/jpet.105.092320. PMID 16326923.
^ Da Settimo F, Taliani S, Trincavelli ML, Montali M, Martini C (2007). "GABA A/Bz receptor subtypes as targets for selective drugs". Current Medicinal Chemistry. 14 (25): 2680–701. doi:10.2174/092986707782023190. PMID 17979718.
^ Lager E, Nilsson J, Østergaard Nielsen E, Nielsen M, Liljefors T, Sterner O (July 2008). "Affinity of 3-acyl substituted 4-quinolones at the benzodiazepine site of GABA(A) receptors". Bioorganic & Medicinal Chemistry. 16 (14): 6936–48. doi:10.1016/j.bmc.2008.05.049. PMID 18541432.
Further reading
.mw-parser-output .refbegin{font-size:90%;margin-bottom:0.5em}.mw-parser-output .refbegin-hanging-indents>ul{list-style-type:none;margin-left:0}.mw-parser-output .refbegin-hanging-indents>ul>li,.mw-parser-output .refbegin-hanging-indents>dl>dd{margin-left:0;padding-left:3.2em;text-indent:-3.2em;list-style:none}.mw-parser-output .refbegin-100{font-size:100%}
Olsen RW, DeLorey TM (1999). "Chapter 16: GABA and Glycine". In Siegel GJ, Agranoff BW, Fisher SK, Albers RW, Uhler MD. Basic neurochemistry: molecular, cellular, and medical aspects (Sixth ed.). Philadelphia: Lippincott-Raven. ISBN 978-0-397-51820-3.
Olsen RW, Betz H (2005). "Chapter 16: GABA and Glycine". In Siegel GJ, Albers RW, Brady S, Price DD. Basic Neurochemistry: Molecular, Cellular and Medical Aspects (Seventh ed.). Boston: Academic Press. pp. 291–302. ISBN 978-0-12-088397-4.
Uusi-Oukari M, Korpi ER (March 2010). "Regulation of GABA(A) receptor subunit expression by pharmacological agents". Pharmacological Reviews. 62 (1): 97–135. doi:10.1124/pr.109.002063. PMID 20123953.
Rudolph U (2015). Diversity and Functions of GABA Receptors: A Tribute to Hanns Möhler (First ed.). Academic Press, Elsevier. ISBN 978-0-12-802660-1.
External links
Receptors,+GABA-A at the US National Library of Medicine Medical Subject Headings (MeSH)
{}}re



Comments
Post a Comment